
 RSS
RSS


人们可以在两个层面上欣赏一件艺术品。 当人们看到古典希腊人的雕刻效果时,我们可以跨越 2,000 多年的距离 感觉 发自内心地认为他们触摸到了一些美丽的东西,并把它变成了石头。 为了将其简化为生物学,我们的感知映射到我们审美判断的进化景观中的深沟。 就像热带草原猿一样,森林的黑暗以其美丽和威严困扰着我们; 但我们是旧石器时代田园草地和边缘的孩子。 同样,在某种程度上,我们在看到身体美时,甚至在我们思考之前就承认了它。*
另一种欣赏层次则较为狭隘,即你能够意识到技术的独创性、执行的精湛技艺和流畅性。 对美学的这一方面的理解自然局限于那些具有同等技能,或者其技能渴望达到大师水平的人。
阅读约瑟夫·拉扎里迪斯的著作 世界首批农民的遗传结构 您可以在两个层面上进行评估。 结果可以广泛获取,但任何尝试过类似方法的人都可以清楚分析的深度。 这些论文出自 戴维·赖希(David Reich) 实验室有一定的模板,但它们绝对不是按数字绘制的。 对于那些对技术细节感兴趣的人, 您 已可以选用 阅读补充材料.
十年前,从这份预印本中收集到的见解在各类研究人员和“基因组博主”眼中只是一线曙光。 现在的问题是从原始结果回到产生结果的动态。 一个深刻的推理问题。
为了达到我们现在的水平,以及大量令人尴尬的结论,研究人员需要做三件事:
1)大量遗传数据以及旨在利用这些数据的方法(基本上是基因组学和用于分析大数据集的统计遗传学)。
2)来自人类过去时间点的遗传数据,而不仅仅是现在。
3) 处理数据所需的技术基础设施(从计算能力到古代 DNA 实验室的神秘艺术)。

我们学到了什么? 古代 DNA 表明,人类过去的遗传变异在时间和空间上都具有很强的不连续性。 这是什么意思呢?
作为一种程式化的事实,在某些方面,将人类变异描述为压倒性的临床变异已成为一种时尚。 也就是说,基因频率作为空间函数的连续变化。 一个相关的事实是预期基因频率会随着时间的推移以类似的稳定和有规律的方式发生变化。
显然,我们物种的临床变异有一定道理。 例如,如果你从法国步行到旁遮普省,那么就很难在基因存在明显不连续性的地方建立一条严格而快速的路线。 但可能会有候选人。 特别是,在中亚的某些地区,您会发现东亚更典型的等位基因频率相当高,而在阿富汗,通常在南亚发现的另一种非西欧亚民族的遗传特征将开始出现。突然出现。
但这两个不连续点说明了一个普遍原则:不连续性是由特定的历史人口事件产生的。 就中亚东亚相关基因比例相当高的情况而言,这几乎可以肯定是东亚相关基因的产物。 突厥扩张,在 1,000 年至 500 年间的约 1500 年中发生过。在南亚,我们现在怀疑有一个相对 最近西欧亚人群的入侵,以及土著群体和收入者之间可能存在一些相互的基因流动。
这两个例子指出,基因流动的重大破坏可能与文化的重大破坏有关。 突厥人的扩张是在历史时期发生的,因此我们可以考察一下,中亚地区伊朗人口的减少始于萨珊王朝晚期,但在700年后蒙古时期的重大冲击中接近完成。 这些事件是 地缘政治 注意。
考虑这一点很重要,因为假设临床变异的旧模型假设遗传变化是通过“群体行动”过程发生的,在这个过程中,小家庭或村庄群体进入了人口扩张阶段,并在字面上超越了其他群体。 这在某种程度上是新石器时代生活方式从近东扩展到欧洲的“人口扩散”理论中隐含的模型,由 Colin Renfew 首创,并由 LL Cavalli-Sforza 及其同事扩展。
在古典经济框架中,人们可以简单地假设那些实行农业生活方式的人将处于边境土地过剩的状态。 因此,他们会有大家庭,并不断扩大他们的活动范围。 以这种方式个人决定 经济人 可以推动大片地区的文化和人口变化 相对 短时间段。

许多人以不协调的方式做出的决定可能会导致我们在周围看到的有序模式,其中包括变异的路线以及基因扩张的信号。 正如 LL Cavalli-Sforza 所指出的,这里的论点并不是说,当使用更新世群体作为土著参考时,现代欧洲人的大多数血统都是外来于欧洲大陆的,而是人口进步的浪潮导致了农业,而不是文化竞争。 即使有了这一波在群体遗传学中得到广泛探索的先进模型,对边境土著群体的同化也意味着,在这一过程结束时,边境上的大多数祖先可能是“土著”。
卡瓦利-斯福尔扎的断言是在 2000 年代初期的一系列结果之后提出的,这些结果被解释为现代欧洲人的大部分祖先来自更新世期间的居民。 这些结果表明农业一定是通过文化扩散而不是人口扩张而传播的。 卡瓦利-斯福尔扎所指出的只是他所支持的模型是关于一个动态过程,而不是按区域进行单倍型计数的某些特定值。
最终这种后卫的道歉是没有必要的。 事实证明,现代欧洲人的大部分血统很可能是过去一万年里外生于欧洲大陆的。 早期用于支持相反结果的结果是正确的,但被误解了。 此外,我还认为卡瓦利-斯福尔扎和他的同事概述的模型在某些方面过于优雅和程式化而没有用处。 如果你读过 文明之前的战争 有大量考古证据表明,史前时期存在大规模的群体间冲突,农民到达非洲大陆可能表现出一些村庄以外的协调和集体行动。 这 波罗的海战役已有 3,200 年历史 这可能是欧洲乃至世界集体行动和冲突的悠久传统的延续。
这是一个“问题”,因为就一般模型而言,地缘政治规模上的群体间冲突不像内生增长参数占主导地位的“前进浪潮”人口情景那样容易处理。 相反,人口统计模式并不是由连续的、可预测的动态造成的, 但这些参数和偶然事件的交集。 历史没有保证,尽管它的车轮倾向于某些有利的凹槽。
二十年前,LL Cavalli-Sforza 写了一本面向非专业读者的书, 伟大的人类流散症。 它是我们一生工作的巅峰之作,它利用源自当代人群的经典标记,调查了我们当时对人类遗传变异的了解。 我们今天拥有的工具要精确得多,拥有数十万个而不是数百个标记,以及来自数千或数万年前人口的 DNA 样本。 研究人员现在不再简单地推断生命之树,而是构建一个关系网格,不仅源自今天可见的节点,而且还源自过去的网格内的位置。
回来的证据是,前现代人群即使在很短的距离内也表现出大量的遗传分化,而且这种分化可能会持续数千年。 群体之间的差异比例约为总差异的 10%,你在欧洲人和汉人之间看到的情况对于附近的民族来说并不罕见,尽管他们之间每代的一个移民会在短时间内消除这种差异。 这种差异平衡最终会被激进的人口更替所打破,因为当地人口灭绝或被新来者吸收,新来者通过扩张重塑了整个景观。 换句话说,如果卡瓦利-斯福尔扎今天要写一本书,我相信它的标题将是“伟大的人类破坏及其离散”。
这不仅仅涉及农业。 来自更新世欧洲的古代DNA 也表明那里的营业额。 欧亚大陆现代人类活动范围的边缘可能存在着某种复合种群动态。 当当地人口灭绝时,新人口就会扩张并占领他们的领土。 古代人类景观可能人口相对稀少,减少了基因流动的机会。
但这可能不是故事的全部。 族群间的冲突无疑发挥了一定作用,古代 DNA 已经发现了相邻族群之间长期存在遗传独特性的证据。 这表明文化习俗成为基因流动的障碍。 今天确实有一个例子发生了这种情况:印度。 种姓制度使得同一地区的当地人群中可以发现大陆上广泛的遗传距离,这些人群已经共存了数千年。
那么拉扎里迪斯论文的结果是什么呢? 顶部的图为您提供了以 PCA 为中心的视图。 基本上,今天所有西欧亚人口都可以被初步近似地建模为四个祖先群体的混合体,这些祖先群体大约在 10,000 年前蓬勃发展。 如果现代遗传变异可以被视为代数,那么对于欧亚大陆西部来说,这就是产生任何合理输出所需的具有不同权重的四个变量。
这四个是:
1)西方狩猎采集者(WHG),欧洲及周边地区的土著居民。
2)东方狩猎采集者(EHG),欧洲东北边缘的土著居民。
3)西方农民,早期欧洲农民(EEF)的祖先,根源于从黎凡特南部向北到安纳托利亚的地区。
4)东方农民,他们是扎根于伊朗西部扎格罗斯山区的人口(中亚农民,CAF)。
这四个人本身就表现出某种复合血统。 EEF 和 CAF 的血统中约有一半是基础欧亚人种 (BEu),这一群体似乎在 50,000 万多年前(即尼安德特人混合之前)就与其他非撒哈拉以南非洲人种分化了。 需要明确的是,BEu 似乎是更新世欧洲狩猎采集者、澳大利亚土著群体和安达曼群岛居民等不同人群的外群体。 EEF 和 CAF 血统的另一半来自两个不同的来源,这解释了它们在 PCA 图上的不同位置。 EEF 具有类似 WHG 的混合物。 也就是说,他们的一些祖先属于更广泛的分支,其中包括欧洲狩猎采集者,以及更遥远的北欧亚祖先(ANE)。 更新世基因组学研究表明,大约 15,000 年前,欧洲狩猎采集者和近东人之间的亲和力大幅增加,这表明存在着将这两个地区联合起来的主要基因流。 这一运动的近东元素可能与 BEu 融合在一起。
其次,CAF 群体的样本数量要少得多,但它似乎与 EHG 有很多共同的祖先,因此两者必定具有来自相关群体的共同祖先。 看来最有可能的来源是 ANE。 由于ANE和WHG之间的遗传距离,EEF和CAF之间的Fst约为0.10,类似于今天中国人和欧洲人之间的Fst。 这两个群体似乎在相似的时间偶然发现了彼此非常接近的农业。
他们在哪里独立事件? 我怀疑他们不是。 我在这里并不是指文化传播。 有证据表明扎格罗斯地区有地方品种的独立驯化。 相反,这两个种群是相似生态景观中更广泛的贸易联系网络的一部分。 两人偶然接触农业并非偶然。 两种相似的农业文化先驱之间很可能存在传播。 它们的位置如此接近不可能是巧合,但具体细节还有待商定。
有趣的是,一旦这两个人口偶然进入农业,他们就向相反的方向扩张。 为什么? 可能是因为他们可以。 也就是说,它们都具有很高的人口密度和社会复杂性,并且没有侵犯彼此的领土,而是扩展到“空旷”的景观。 狩猎采集者居住的地区更容易被消灭或同化。 卡迪尔人和LBK人在欧洲的传播速度如此之快,几乎可以肯定他们最初都是一个文化单位。 类似的情况可能也适用于向东扩展到南亚、向北扩展到草原的非洲武装部队团体。
本文的另一个有趣的结果是,WHG 本身似乎是东部人群的混合物。 更准确地说,早期分析中用作 WHG“纯粹”范例的中石器时代狩猎采集者,结果却并非如此,而是表现出与其他群体的某种混合。 这可能就是本文中 EHG 的 ANE 比例要高得多的原因。 来自瑞士比雄犬的较旧样本缺乏东方混合物,因此可以作为 WHG 的更好参考。 尽管还不确定,但现在看来,ANE 与东欧亚人(例如汉族)的混合导致了这些人群与当今欧洲人之间的某种亲和力,这可以追溯到但不限于 WHG。 这并不奇怪。 农业的出现并不是什么新鲜事,无论何时何地,文化创新似乎都会引发人口混乱。
尽管本预印本的核心内容是四个种群足以解释遗传变异,并且 西欧亚人口的人口历史, 我认为也许更有趣的元素是 ANE 和 BEu 的角色。 如今,这两个群体都不以“纯粹”的形式存在。 我们不知道 BEu 是谁。 我们不知道他们从哪里来。 在我看来,BEu 血统在 EEF(至少是它们在中东的前身)和 CAF 中的比例大致相同,这一点值得怀疑。 这两个 BEu 组件似乎并没有太大区别。 对我来说,这表明 BEu 最近可能有所扩张。 我还相信 BEu 可能在调节“回到非洲”基因流中发挥作用。 由于 BEu 缺乏尼安德特人的混合物,这可以解释该大陆大部分地区的 BEu 水平非常低,但目前看来是欧亚起源的 EY 染色体单倍型的存在。
至于 ANE,其地理覆盖范围令人难以置信,从西欧一直到新世界。 看起来,作为一个不混合的群体,他们一直坚持到全新世,但在数量上他们总是捉襟见肘。 通过与农业学家的融合,他们得以保留下来,但他们的许多古西伯利亚民俗可能已经消失。 我确实相信 Y 染色体上的 R1 单倍群很可能源自它们,因为它是美洲印第安人 Q 的姐妹。
报纸上有很多东西值得咀嚼,尤其是补充材料。 例如,“草原”血统在整个南亚所占的比例并不小。 这该怎么办? 我想我会推迟到古代 DNA 出现为止,就像接下来的 6 个月一样。
但是,我确实认为古代 DNA 和模型 破坏和不连续性支持了这样一个命题:间断平衡作为一个论点对文化进化比对生物学更有效。 文化表现出惯性和从众倾向。 学习新事物很困难。 随着狩猎采集者从兼性耕种转变为强制农作物生产方式,农业在近东的“占领”必须存在非常特殊的条件。 一旦这些文化成为农耕文化,邻居们就不容易轻易采用它们,因为文化包往往是一个整体,包含许多偶然的部分。 农业的优势在于它可以从土地中获取更多的产量,并且人口密度也会增加。 更高的人口密度意味着在群体间冲突中拥有更多资源(如果涉及到这一点),并且需要扩大规模以继续超越马尔萨斯极限。 一旦空间被占据,就会达到新的平衡。
我想重申,这种模式不仅仅适用于农业。 斯宾塞·威尔斯的推文:
巴布亚新几内亚令人着迷——期待看到这篇文章的出版。 #SMBE16 pic.twitter.com/8fhB8Pcaqv
— 斯宾塞·威尔斯 (@spwells) 2016 年 7 月 4 日
新几内亚是一个园艺社会,高地人口非常稠密。 高 Fst 与全新世早期欧亚大陆西部或今天印度的情况一致。 但观察遗传分化是从过去一万年开始的,而不是过去的 50,000。 一万年前是什么时候 园艺 开始了。 很可能高地的一个人口开始实行这一做法,并扩大人口规模,消灭或吸收其邻居。 但新几内亚的地形对可能性的范围设置了严格的限制,因为该岛的高地是一个非常孤立的生态系统。 一旦扩张阶段结束,遗传分化就开始了,群体开始在马尔萨斯极限下为生存而斗争。
Lazaridis 等人的论文的见解之一是,这种情况并未发生在欧亚大陆。 EEF 和 CAF 之间的差异逐渐缩小,因为近东在青铜时代见证了相互的基因流动。 差异不在于农业,而在于后农业社会的复杂性,这使得彼得·图尔钦(Peter Turchin)所谓的“元种族”身份和超越地域的复杂机构的出现成为可能。 在新的均衡状态下,Fst 并没有随着人口争夺资源而开始上升,因为创新开始轻轻地将生产边界向外推,并促进物质(例如贸易)和思想(例如宗教)的联系。
整个故事还没有一成不变。 接下来的几年将会很有趣。 中国是下一个前沿,古代DNA很快就会开启它的历史。 但在我们眼前见证史前史的揭开是一个激动人心的时刻。
* 审美判断的一个维度是受文化条件限制的,而另一个维度则不受文化条件限制。 我这里说的是后者。





是的。
关于 y DNA 单倍型 E 的起源,在某些方面是一个非常非常敏感的话题,出于类似的原因,y DNA r1a1a 在某些其他种族中也是敏感的。
坦率地说,E 在非洲占主导地位。如果E最终是欧亚人种(这一点存在激烈争议),那么这意味着在古代欧亚人种占主导地位的非洲人,这一假设对一些人来说很难接受。
然而,目前还没有关于 E 起源于欧亚大陆的明确案例,因此“还没有定论”。
现在说还为时过早。但在更易于管理的时间尺度上,在纳图夫语系中检测到各种 E1b1b 亚群也迫使人们对亚非语系 urheimat 问题进行跨学科重新审视——新的综合是否会像印欧语系那样落后于 CW?
我一直认为贝尔伍德在这里是错误的,因为奥莫提克和库希提克有很大的分歧——但类似纳图夫的基因流(和近东技术)流向北非和东非的信号比相反的要强得多,这使得我想知道。更不用说,E-Z830(其中一些是 E-M123,如果 Genetiker 是正确的话)在以色列的早期存在,使闪米特语作为亚非语系单一的、晚期亚洲分支的概念变得严重复杂化。
话又说回来,亚非语系(甚至是闪米特语系)的年代范围是如此之深,而非洲-黎凡特联系的本质又如此潜在地错综复杂,以至于古代DNA可能远不如IE的情况那样具有决定性。不过,至少,这对于 E 的故事来说是有帮助的。
回复:@Anonymous
我认为拉齐布说“欧亚”时指的是 Y-DNA E-M215 (E1b1b) 和/或 E-M35(E1b1b 的后代)。 E-M35 主要发现于具有显着或大部分西欧亚血统的人群(有角非洲人、西北非洲人、埃及人、希腊人等)。坦率地说,我怀疑它的起源是在西亚,但是是的。你指的是拉齐布的“E”吗?
我认为您可能已经知道一些注释,但可能会为其他读者澄清:
1)在集群与克隆方面,安纳托利亚新石器时代版本的 EEF 与黎凡特新石器时代版本的 EEG 相比,与欧洲 WHG 和高加索 HG 的相对相关性确实表明,古代中东和近东地区存在临床变异。至少到了新石器时代早期/中石器时代。
WHG 内部似乎也有类似的结构,我们所掌握的匈牙利 HG 样本与 EHG 和安纳托利亚的差异似乎小于来自西欧相似时期的 WHG(尽管这在统计上似乎仍然相当微妙)。
因此,看起来它确实可能在相关性的空间结构中具有临床意义,尽管相对于今天而言,这些细胞系内的种群内的分化非常低(因此 FST 差异非常高)。看来,遗传结构在地理上是否具有临床性的问题,与相对于距离比今天更高或更低的遗传分化是不同的?
2) 我不知道 EEF 和 CAF 之间的 fst 是否约为 0.100。论文补充中给出的 fsts 包括:
黎凡特N-伊朗_N : 0.047
安纳托利亚_N-伊朗_N : 0.071
Europe_EN-Iran_N:0.071
这比最近的欧洲汉姆稍低:0.110。
LevantN-Han:0.139,Iran_N-Han:0.123,Anatolia_N-Han:0.133,Europe_NE-Han:0.129
同时,WHG-Iran_N:0.126 以及 WHG-Han:0.162
因此,新石器时代早期近东群体之间的分化可能比当今欧洲人和汉族之间的分化强 0.5,而 WHG 与其最不相似的中东亲戚之间的分化比当今欧洲人和汉族之间的分化强,但需要注意的是,WHG-汉族分化也比当今欧洲人和汉族之间的分化更强。对于当今的欧洲人来说。
Fst 似乎对这里的某些东西表示怀疑,可能是数据集大小的变化。
Granot 等人在法国-日本获得了 0.085,并且在可比统计数据上的距离似乎比 Lazaridis 2016 更短。 他们使用了 660k SNP。
http://biorxiv.org/content/biorxiv/suppl/2016/05/25/033852.DC2/033852-1.pdf
从 Lazaridis 论文的表 S4 中不清楚有多少 SNP 用于区分 Fst。 如果它与 ADMIXTURE 分析和一些正式测试中的相同,则可能更接近 250k。
无论如何,看到 Lazaridis 集的 Est 中位数会很棒。
回复:@M
1)在集群与克隆方面,安纳托利亚新石器时代版本的 EEF 与黎凡特新石器时代版本的 EEG 相比,与欧洲 WHG 和高加索 HG 的相对相关性确实表明,古代中东和近东地区存在临床变异。至少到了新石器时代早期/中石器时代。
WHG 内部似乎也有类似的结构,我们所掌握的匈牙利 HG 样本与 EHG 和安纳托利亚的差异似乎小于来自西欧相似时期的 WHG(尽管这在统计上似乎仍然相当微妙)。
因此,看起来它确实可能在相关性的空间结构中具有临床意义,尽管相对于今天而言,这些细胞系内的种群内的分化非常低(因此 FST 差异非常高)。看来,遗传结构在地理上是否具有临床性的问题,与相对于距离比今天更高或更低的遗传分化是不同的?
2) 我不知道 EEF 和 CAF 之间的 fst 是否约为 0.100。论文补充中给出的 fsts 包括:
黎凡特N-伊朗_N : 0.047
安纳托利亚_N-伊朗_N : 0.071
Europe_EN-Iran_N:0.071
这比最近的欧洲汉姆稍低:0.110。
LevantN-Han:0.139,Iran_N-Han:0.123,Anatolia_N-Han:0.133,Europe_NE-Han:0.129
同时,WHG-Iran_N:0.126 以及 WHG-Han:0.162
因此,新石器时代早期近东群体之间的分化可能比当今欧洲人和汉族之间的分化强 0.5,而 WHG 与其最不相似的中东亲戚之间的分化比当今欧洲人和汉族之间的分化强,但需要注意的是,WHG-汉族分化也比当今欧洲人和汉族之间的分化更强。对于当今的欧洲人来说。
回复:@Shaikorth
Fst 似乎对这里的某些东西表示怀疑,可能是数据集大小的变化。
Granot 等人的法语-日语得分为 0.085,并且与 Lazaridis 2016 相比,可比统计数据的距离通常更短。他们使用了 660k SNP。
http://biorxiv.org/content/biorxiv/suppl/2016/05/25/033852.DC2/033852-1.pdf
从 Lazaridis 论文的表 S4 中并不清楚有多少 SNP 用于区分 Fst。如果与 ADMIXTURE 分析和一些正式测试中的相同,则可能接近 250k。
无论如何,看到 Lazaridis 集的 Est 中位数会很棒。
我认为这听起来像是一个极端异常的数字 IRC。尽管如此,我不太热衷于比较不同 fst 矩阵的 fst 距离,或者说总体 x 和 y 之间的 fst *是*一个特定值(从一个矩阵到另一个矩阵存在可变性)。
IRC fsts 从一个矩阵到另一个矩阵往往相对相同,但实际数字往往会有所不同,例如 Granot:Italian-Yoruba = 0.115,French-Japanese = 0.085,Laziridis:Italian-Yoruba = 0.148,French-Han = 0.128 . 可比对的比率约为 0.66-0.77。 不过,它可能更一致,所以这是有问题的原因。
不知道 est 对样本大小是否或多或少敏感。 IRC那篇论文建议比较敏感。
关于 y DNA 单倍型 E 的起源,在某些方面是一个非常敏感的话题,出于类似的原因,y DNA r1a1a 在某些其他种族中也是敏感的。
坦率地说,E 在非洲占主导地位。如果E最终是欧亚人种(这一点存在激烈争议),则意味着在古代欧亚人种占主导地位的非洲人,这一假设对一些人来说很难接受。
然而,目前还没有关于 E 起源于欧亚大陆的明确案例,因此“还没有定论”。
回复:@ben-canaan、@Awale
现在说还为时过早。但在更易于管理的时间尺度上,在纳图夫语系中检测到各种 E1b1b 亚群也迫使人们对亚非语系的 urheimat 问题进行跨学科重新审视——新的综合是否会像印欧语系那样落后于 CW?
我一直认为贝尔伍德在这里是错误的,因为奥莫提克和库希提克的巨大分歧——但北非和东非的类似纳图夫基因流(和近东技术)的信号比相反的要强得多,这让我想知道。更不用说,E-Z830(其中一些是 E-M123,如果 Genetiker 是正确的话)在以色列的早期存在,使闪米特语作为亚非语系单一的、晚期亚洲分支的概念变得严重复杂化。
话又说回来,亚非语系(甚至是闪米特语系)的年代范围是如此之深,而非洲-黎凡特联系的本质又如此潜在地错综复杂,以至于古代DNA可能远不如IE的情况那样具有决定性。不过,至少,这对于 E 的故事来说是有帮助的。
作为一个外行,我再补充两分钱。
单倍型 E 与单倍型 D 形成一个分支。如今,单倍型 D 集中在东亚人中,尤其是日本人中。
也许基础欧亚DE人群生活在非常遥远的时代,以至于东亚人和黎凡特人类型之间的特征性表型差异甚至还没有形成。
回复:@Razib Khan
他们扩展到南亚,但并没有真正进入草原。单亲不匹配。
他们迁入南亚后,他们的祖国大部分被从西方迁入的新人口所取代,从而形成了伊朗西部的红铜时代人口。
这个种群的祖先很可能也通过高加索地区迁入了草原,在那里他们获得了 CHG 混合物,然后与 EHG 混合。
但没有人知道现阶段它们来自哪里,以及它们到底为草原牧民做出了多少贡献。也许他们是来自黎凡特北部的哈拉夫牧民?
Iosif 不知道,但他目前将 Steppe_EMBA 建模为 57% EHG,43% 伊朗铜石。工作正在进行中。
Karmin 等人发现,Y-DNA 多样性的瓶颈可追溯到最初的新石器时代之后。等人。发现近东地区从红铜时代到青铜时代常染色体组成的快速而连续的变化存在良好的相关性。团体间的竞争一定非常激烈。
希望他们对新石器时代后基因组中的选择迹象进行后续研究,因为我们现在有伊朗霍图河和甘吉达雷河。
我在这篇新论文中一直在考虑的一件事是,近东和东亚采用农业的动态现在似乎是相似的。我们知道,在东亚,至少有许多不同的语言群体——汉藏语系、南亚语系、南岛语系、壮族语系和苗族语系——似乎都起源于现在的中国北部或中部地区。此外,IIRC 即将发表的一篇论文表明,韩国人和日本人的祖先是狩猎采集者,他们基本上从事农业,没有任何来自中国北方的主要外来混合物。因此,东亚的总体模式似乎是许多不同的群体在新石器时代早期独立从事农业,后来扩散到周围的“空旷”地区。这似乎与西欧亚大陆不同,人们认为那里有一个人口在功能上席卷了一切,并与狩猎采集者混合在一起——因为我们知道这就是向欧洲和南亚扩张的动力。
当然,这篇论文表明,至少有 2-3 个不同的种群最初在近东从事农业,随着时间的推移,它们在基因上越来越接近,直到形成一个“种族”。鉴于近东最早记录的语言之间存在显着的语言多样性,我们确实应该猜到这一点。我的理解是,更多的古代DNA论文正在从东亚运来,所以我想我们很快就会发现更多——这很好,因为这似乎是现代西欧亚人种基础的主要谜团(除了基础欧亚人种之外)是)现已解决。
巴布亚的论文看起来很有趣,而且结果老实说并不太令人惊讶。从我读到的和从照片中看到的情况来看,巴布亚人表现出非常广泛的表型多样性——甚至不考虑最近的南岛语混合物。有些人很容易被误认为撒哈拉以南非洲人,而另一些人的面部特征几乎是西欧亚人——以至于如果他们的头发不卷曲,他们在南亚就不会显得格格不入。 (本尼文达就是一个很好的例子)。身高和肤色的差异似乎也因种族而有很大差异。因此,如果这种表型多样性反映在基因型中也就不足为奇了。
顺便说一句,我最近读到,基于 DNA 确定面部结构的方法已经有了显着改进。如此之多,以至于人们可以使用这些面部模拟从队列中正确地挑选出个人,这种情况的成功率高达 90%。古代 DNA 的覆盖范围是否足够好,以至于我们可以用这种方式重建古代面孔?我认为它作为《发现》或《国家地理》等科普杂志上的一篇文章会受到爆炸性的欢迎。
该论文发现,9 年前东北亚的类似乙支的狩猎采集人群与日本人和韩国人有相似之处,其基因组的其余部分被建模为最接近华南少数民族或东南亚人,这是有道理的,因为新石器时代的包韩国和日本以稻米为中心,与中国南方新石器时代相同,与华北平原的小米不同。因此,就新石器时代而言,人口与技术之间的紧密联系仍然一如既往地紧密。
汉藏语年龄不超过6-7年前,南岛语则稍年轻一些。东南亚多个地区的南亚农民被后来的南岛人套印,通常是在原始历史时期。壮侗语似乎是南岛语系回迁到大陆的语言,创造了一个具有广泛的再词汇化的超级语言,它们的传播也很晚,在湄公河以南地区的历史时期。朝鲜语似乎侵入了朝鲜半岛,而朝鲜半岛似乎最早就居住着说日语的人。因此,亚洲绝对不是新石器时代后基因流动和文化搅动模式的例外,而且那里的遗传景观特征不太可能在最早的新石器时代就已确立。
回复:@Karl Zimmerman
除其他外,Lazaridis 2016 的补充包括一个模型,该模型将东亚集合种群拟合为“古代南欧亚人”(类似翁格)和古代北欧亚人(类似马耳他)的某种混合体,就像美洲原住民一样。如果模型成立,那将是一件大事。问题之一可能是马耳他现在对 Onge 的亲和力超出了 EHG 和 WHG 的水平。
我预计巴布亚人的遗传状况以自己的方式反映了美洲原住民的遗传状况——不同人群之间存在一系列表型多样性和高 Fst,但总体上都具有相同的祖先群体组成。
回复:@Awale、@Karl Zimmerman
向往大师的境界
不止优秀!
优秀的帖子。 Seomwhat 是技术性的,但服务于将基因组学纳入(前)历史的角度。很喜欢这个!
关于 y DNA 单倍型 E 的起源,在某些方面是一个非常敏感的话题,出于类似的原因,y DNA r1a1a 在某些其他种族中也是敏感的。
坦率地说,E 在非洲占主导地位。如果E最终是欧亚人种(这一点存在激烈争议),则意味着在古代欧亚人种占主导地位的非洲人,这一假设对一些人来说很难接受。
然而,目前还没有关于 E 起源于欧亚大陆的明确案例,因此“还没有定论”。
回复:@ben-canaan、@Awale
我认为拉齐布说“欧亚”时指的是 Y-DNA E-M215 (E1b1b) 和/或 E-M35(E1b1b 的后代)。 E-M35 主要发现于具有显着或大部分西欧亚血统的人群(有角非洲人、西北部非洲人、埃及人、希腊人等)。坦率地说,我怀疑它的起源是在西亚,但是是的。你指的是拉齐布的“E”吗?
当然,这篇论文表明,至少有 2-3 个不同的种群最初在近东从事农业,随着时间的推移,它们在基因上越来越接近,直到形成一个“种族”。鉴于近东最早记录的语言之间存在显着的语言多样性,我们确实应该猜到这一点。我的理解是,更多的古代DNA论文正在从东亚运来,所以我想我们很快就会发现更多——这很好,因为这似乎是现代西欧亚人种基础的主要谜团(除了基础欧亚人种之外)是)现已解决。
巴布亚的论文看起来很有趣,而且结果老实说并不太令人惊讶。从我读到的和从照片中看到的情况来看,巴布亚人表现出非常广泛的表型多样性——甚至不考虑最近的南岛语混合物。有些人很容易被误认为是撒哈拉以南非洲人,而另一些人的面部特征几乎是西欧亚人——以至于如果他们的头发不卷曲,他们在南亚就不会显得格格不入。 (本尼文达就是一个很好的例子)。身高和肤色的差异似乎也因种族而有很大差异。因此,如果这种表型多样性反映在基因型中也就不足为奇了。
顺便说一句,我最近读到,基于 DNA 确定面部结构的方法已经有了显着改进。如此之多,以至于人们可以使用这些面部模拟从队列中正确地挑选出个人,这种情况的成功率高达 90%。古代 DNA 的覆盖范围是否足够好,以至于我们可以用这种方式重建古代面孔?我认为它作为《发现》或《国家地理》等科普杂志上的一篇文章会受到爆炸性的欢迎。
回复:@RK、@Shaikorth
该论文发现,9 年前东北亚的类似乙支的狩猎采集人群与日本人和韩国人有相似之处,其基因组的其余部分被建模为最接近华南少数民族或东南亚人,这是有道理的,因为新石器时代的包韩国和日本以稻米为中心,与中国南方新石器时代相同,与华北平原的小米不同。因此,就新石器时代而言,人口与技术之间的紧密联系仍然一如既往地紧密。
汉藏语年龄不超过6-7年前,南岛语则稍年轻一些。东南亚多个地区的南亚农民被后来的南岛人套印,通常是在原始历史时期。壮侗语似乎是南岛语系回迁到大陆的语言,创造了一个具有广泛的再词汇化的超级语言,它们的传播也很晚,在湄公河以南地区的历史时期。朝鲜语似乎侵入了朝鲜半岛,而朝鲜半岛似乎最早就居住着说日语的人。因此,亚洲绝对不是新石器时代后基因流动和文化搅动模式的例外,而且那里的遗传景观特征不太可能在最早的新石器时代就已确立。
感谢您对最近论文的更正...我正在四处寻找它,但我什至不记得作者,所以我没有太多运气来刷新自己的细节。
我并不是说东亚没有基因搅动——只是说,在新石器时代,许多不同的群体似乎独立地从事农业,其中许多人以某种形式幸存至今。近东新石器时代似乎也存在同样的普遍模式,但随着时间的推移,遗传和语言差异往往会稳步减少。我怀疑奴隶制的发展在这方面发挥了重要作用——在中石器文化中,俘虏的敌人在经济上毫无用处,但额外的劳动力对农民来说总是有用的。因此,技术复杂程度大致相似的两种文化如果彼此共处,可能会定期互相袭击,以吸引妇女和儿童来增加劳动力,从而在基因型和表型(也许不是文化)方面慢慢地将人口编织在一起。
当然,这篇论文表明,至少有 2-3 个不同的种群最初在近东从事农业,随着时间的推移,它们在基因上越来越接近,直到形成一个“种族”。鉴于近东最早记录的语言之间存在显着的语言多样性,我们确实应该猜到这一点。我的理解是,更多的古代DNA论文正在从东亚运来,所以我想我们很快就会发现更多——这很好,因为这似乎是现代西欧亚人种基础的主要谜团(除了基础欧亚人种之外)是)现已解决。
巴布亚的论文看起来很有趣,而且结果老实说并不太令人惊讶。从我读到的和从照片中看到的情况来看,巴布亚人表现出非常广泛的表型多样性——甚至不考虑最近的南岛语混合物。有些人很容易被误认为是撒哈拉以南非洲人,而另一些人的面部特征几乎是西欧亚人——以至于如果他们的头发不卷曲,他们在南亚就不会显得格格不入。 (本尼文达就是一个很好的例子)。身高和肤色的差异似乎也因种族而有很大差异。因此,如果这种表型多样性反映在基因型中也就不足为奇了。
顺便说一句,我最近读到,基于 DNA 确定面部结构的方法已经有了显着改进。如此之多,以至于人们可以使用这些面部模拟从队列中正确地挑选出个人,这种情况的成功率高达 90%。古代 DNA 的覆盖范围是否足够好,以至于我们可以用这种方式重建古代面孔?我认为它作为《发现》或《国家地理》等科普杂志上的一篇文章会受到爆炸性的欢迎。
回复:@RK、@Shaikorth
除其他外,Lazaridis 2016 的补充包括一个模型,该模型将东亚集合种群拟合为“古代南欧亚人”(类似翁格)和古代北欧亚人(类似马耳他)之间的某种混合体,就像美洲原住民一样。如果模型成立,那将是一件大事。问题之一可能是马耳他现在对 Onge 的亲和力超出了 EHG 和 WHG 的水平。
我预计巴布亚人的遗传状况以自己的方式反映了美洲原住民的遗传状况——不同人群之间存在一系列表型多样性和较高的 Fst,但总体上都具有相同的祖先群体组成。
您不认为 David 的树混合有点与“Han = ANE-like + Onge-like”模型相矛盾吗?他们让所有东部非非洲人都对 ANE 表现出一种奇怪的亲和力:
https://drive.google.com/file/d/0B9o3EYTdM8lQM1BINGE3V2hJczA/view
https://drive.google.com/file/d/0B9o3EYTdM8lQZGp5VWd5M0gtTkU/view
?
这是否基本上意味着美洲原住民获得了“双倍剂量”的马耳他——一剂来自原始东亚人群,另一剂来自美洲人的种族形成?或者这是否意味着在原始美洲人形成时,还没有类似东亚人的东西存在?
如果是后者,这是否有助于解释一个假设,即有证据表明,OOA 很早就移民到了亚洲,这在今天的奥美拉尼西亚血统中得到了体现?也许除奥美拉尼西亚人以外的所有欧亚人都具有 ANE 混合物,导致后者看起来比实际情况更基础。
回复:@Shaikorth
除其他外,Lazaridis 2016 的补充包括一个模型,该模型将东亚集合种群拟合为“古代南欧亚人”(类似翁格)和古代北欧亚人(类似马耳他)的某种混合体,就像美洲原住民一样。如果模型成立,那将是一件大事。问题之一可能是马耳他现在对 Onge 的亲和力超出了 EHG 和 WHG 的水平。
我预计巴布亚人的遗传状况以自己的方式反映了美洲原住民的遗传状况——不同人群之间存在一系列表型多样性和高 Fst,但总体上都具有相同的祖先群体组成。
回复:@Awale、@Karl Zimmerman
您不认为 David 的树混合有点与“Han = ANE-like + Onge-like”模型相矛盾吗?他们让所有东部非非洲人都对 ANE 表现出一种奇怪的亲和力:
https://drive.google.com/file/d/0B9o3EYTdM8lQM1BINGE3V2hJczA/view
https://drive.google.com/file/d/0B9o3EYTdM8lQZGp5VWd5M0gtTkU/view
?
该论文发现,9 年前东北亚的类似乙支的狩猎采集人群与日本人和韩国人有相似之处,其基因组的其余部分被建模为最接近华南少数民族或东南亚人,这是有道理的,因为新石器时代的包韩国和日本以稻米为中心,与中国南方新石器时代相同,与华北平原的小米不同。因此,就新石器时代而言,人口与技术之间的紧密联系仍然一如既往地紧密。
汉藏语年龄不超过6-7年前,南岛语则稍年轻一些。东南亚多个地区的南亚农民被后来的南岛人套印,通常是在原始历史时期。壮侗语似乎是南岛语系回迁到大陆的语言,创造了一个具有广泛的再词汇化的超级语言,它们的传播也很晚,在湄公河以南地区的历史时期。朝鲜语似乎侵入了朝鲜半岛,而朝鲜半岛似乎最早就居住着说日语的人。因此,亚洲绝对不是新石器时代后基因流动和文化搅动模式的例外,而且那里的遗传景观特征不太可能在最早的新石器时代就已确立。
回复:@Karl Zimmerman
感谢您对最近论文的更正……我正在四处寻找它,但我什至不记得作者,所以我没有太多运气来刷新自己的细节。
我并不是说东亚没有基因搅动——只是说,在新石器时代,许多不同的群体似乎独立地从事农业,其中许多人以某种形式幸存至今。近东新石器时代似乎也存在同样的普遍模式,但随着时间的推移,遗传和语言差异往往会稳步减少。我怀疑奴隶制的发展在其中发挥了重要作用——在中石器文化中,被俘虏的敌人在经济上毫无用处,但额外的劳动力对农民来说总是有用的。因此,技术复杂程度大致相似的两种文化如果彼此共处,可能会定期互相袭击,以吸引妇女和儿童来增加劳动力,从而在基因型和表型(也许不是文化)方面慢慢地将人口编织在一起。
看起来 ANE 和 Onge 确实有着欧洲 HG 所没有的密切关系。然后,在伊比利亚新石器时代和 LBK 的树中,在 Papuan/Onge 分裂后,Dai/Han/Karitiana 根与 ANE 之间有超过 20% 的边缘(然后是 Karitiana 的另一个边缘)。
在另一组树中,ryukendo 询问 Onge 分裂后,东亚人(但不是获得特定 ANE 的美洲原住民)和欧洲 HG/ANE 分支之间存在 16% 的优势。 我不会说树木排除了东亚人的 ASE+ANE 模型,但 Onge 或 ANE 本身可能有一段复杂的历史……
哦,顺便说一句,我的错......我最初误读了我发送的一些树混合。
在另一组树中,ryukendo 询问 Onge 分裂后,东亚人(但不是获得特定 ANE 的美洲原住民)和欧洲 HG/ANE 分支之间存在 16% 的优势。 我不会说树木排除了东亚人的 ASE+ANE 模型,但 Onge 或 ANE 本身可能有一段复杂的历史……"
是的,我想我们最终需要更多的数据和古代基因组(比如来自南亚和东亚的基因组)。 当我们拥有这些样本时,看看会发生什么应该很有趣。
回复:@PQ
现在说还为时过早。但在更易于管理的时间尺度上,在纳图夫语系中检测到各种 E1b1b 亚群也迫使人们对亚非语系 urheimat 问题进行跨学科重新审视——新的综合是否会像印欧语系那样落后于 CW?
我一直认为贝尔伍德在这里是错误的,因为奥莫提克和库希提克有很大的分歧——但类似纳图夫的基因流(和近东技术)流向北非和东非的信号比相反的要强得多,这使得我想知道。更不用说,E-Z830(其中一些是 E-M123,如果 Genetiker 是正确的话)在以色列的早期存在,使闪米特语作为亚非语系单一的、晚期亚洲分支的概念变得严重复杂化。
话又说回来,亚非语系(甚至是闪米特语系)的年代范围是如此之深,而非洲-黎凡特联系的本质又如此潜在地错综复杂,以至于古代DNA可能远不如IE的情况那样具有决定性。不过,至少,这对于 E 的故事来说是有帮助的。
回复:@Anonymous
作为一个外行,我再补充两分钱。
单倍型 E 与单倍型 D 形成一个分支。如今,单倍型 D 集中在东亚人中,尤其是日本人中。
也许基础欧亚DE人群生活在非常遥远的时代,以至于东亚人和黎凡特人类型之间的特征性表型差异甚至还没有形成。
我们看到的表型是过去一万年的产物。所以它肯定早于那个。
除其他外,Lazaridis 2016 的补充包括一个模型,该模型将东亚集合种群拟合为“古代南欧亚人”(类似翁格)和古代北欧亚人(类似马耳他)的某种混合体,就像美洲原住民一样。如果模型成立,那将是一件大事。问题之一可能是马耳他现在对 Onge 的亲和力超出了 EHG 和 WHG 的水平。
我预计巴布亚人的遗传状况以自己的方式反映了美洲原住民的遗传状况——不同人群之间存在一系列表型多样性和高 Fst,但总体上都具有相同的祖先群体组成。
回复:@Awale、@Karl Zimmerman
这是否基本上意味着美洲原住民获得了“双倍”的马耳他——一剂来自原始东亚人群,另一剂来自美洲人的种族形成?或者这是否意味着在原始美洲人形成时,还没有类似东亚人的东西存在?
如果是后者,这是否有助于解释一个假设,即有证据表明,OOA 很早就移民到了亚洲,这在今天的奥美拉尼西亚血统中得到了体现?也许除奥美拉尼西亚人以外的所有欧亚人都具有 ANE 混合物,导致后者看起来比实际情况更基础。
不管怎样,正确的答案都需要来自东亚的古代DNA。正如您提到的,在 Treemix 运行中,Karitiana 获得双倍剂量,有些则获得单边缘,而 Dai/Han 则有单独的边缘。
这是否基本上意味着美洲原住民获得了“双倍剂量”的马耳他——一剂来自原始东亚人群,另一剂来自美洲人的种族形成?或者这是否意味着在原始美洲人形成时,还没有类似东亚人的东西存在?
如果是后者,这是否有助于解释一个假设,即有证据表明,OOA 很早就移民到了亚洲,这在今天的奥美拉尼西亚血统中得到了体现?也许除奥美拉尼西亚人以外的所有欧亚人都具有 ANE 混合物,导致后者看起来比实际情况更基础。
回复:@Shaikorth
不管怎样,正确的答案都需要来自东亚的古代DNA。正如您提到的,在 Treemix 运行中,Karitiana 获得双倍剂量,有些则获得单边缘,而 Dai/Han 则有单独的边缘。
在另一组树中,ryukendo 询问 Onge 分裂后,东亚人(但不是获得特定 ANE 的美洲原住民)和欧洲 HG/ANE 分支之间存在 16% 的优势。我不会说树木排除了东亚人的 ASE+ANE 模型,但 Onge 或 ANE 本身可能有一段复杂的历史......
回复:@Awale
哦,顺便说一句,我的错……我最初误读了我发送的一些树混音。
在另一组树中,ryukendo 询问 Onge 分裂后,东亚人(但不是获得特定 ANE 的美洲原住民)和欧洲 HG/ANE 分支之间存在 16% 的优势。 我不会说树木排除了东亚人的 ASE+ANE 模型,但 Onge 或 ANE 本身可能有一段复杂的历史……=
是的,我想我们最终需要更多的数据和古代基因组(比如来自南亚和东亚的基因组)。 当我们拥有这些样本时,看看会发生什么应该很有趣。
将 ANE 输入到所有 ENA 祖先的模型涉及 WHG 和 ANE,作为来自以伊朗_N 为根基的更大基底混合组的最终提示; 此外,ANE 与 WHG 的边缘共享最多 3% 的漂移,即两者之间根本没有太大区别。 边缘很可能涉及 WHG 和 ANE 被拉近 ENA,因为需要考虑它们的非 Basal 与其他非 Basal 的接近程度。
不排除 ANE 和 ENA 有着复杂的历史; Chad 和我去检查了一些统计数据,其中 WHG-ANE 分支与 ENA 大致对称,但 MA-1 相对接近巴布亚人和 Onge,EHG 与东亚人狭义定义,一些 WHG 与东亚人狭义定义,SHG 不接近去任何; 但是 Treemix 中树的一个部分上的边缘可以是对另一部分上的现象的补偿性调整。
作为一个外行,我再补充两分钱。
单倍型 E 与单倍型 D 形成一个分支。如今,单倍型 D 集中在东亚人中,尤其是日本人中。
也许基础欧亚DE人群生活在非常遥远的时代,以至于东亚人和黎凡特人类型之间的特征性表型差异甚至还没有形成。
回复:@Razib Khan
我们看到的表型是过去一万年的产物。所以它肯定早于那个。
Fst 似乎对这里的某些东西表示怀疑,可能是数据集大小的变化。
Granot 等人在法国-日本获得了 0.085,并且在可比统计数据上的距离似乎比 Lazaridis 2016 更短。 他们使用了 660k SNP。
http://biorxiv.org/content/biorxiv/suppl/2016/05/25/033852.DC2/033852-1.pdf
从 Lazaridis 论文的表 S4 中不清楚有多少 SNP 用于区分 Fst。 如果它与 ADMIXTURE 分析和一些正式测试中的相同,则可能更接近 250k。
无论如何,看到 Lazaridis 集的 Est 中位数会很棒。
回复:@M
我认为这听起来像是一个极端的离群数字 IRC。 虽然,我不太热衷于比较不同 fst 矩阵的 fst 距离,或者说群体 x 和 y 之间的 fst *是* 出于这个原因的某个值(从一个矩阵到另一个矩阵存在可变性)。
IRC fsts 从一个矩阵到另一个矩阵往往相对相同,但实际数字往往会有所不同,例如 Granot:Italian-Yoruba = 0.115,French-Japanese = 0.085,Laziridis:Italian-Yoruba = 0.148,French-Han = 0.128 . 可比对的比率约为 0.66-0.77。 不过,它可能更一致,所以这是有问题的原因。
不知道 est 对样本大小是否或多或少敏感。 IRC那篇论文建议比较敏感。
拉齐卜: 报纸上有很多东西值得咀嚼,尤其是补充材料。
我认为另一件值得一提的事情是黎凡特新石器时代的人(如果不是纳图夫人的话)似乎是东非西欧亚相关血统的原因。
我的观点是,这是一个很好的发现,因为它基本上协调了基于正式统计的证据,这些证据往往认为 EEF 是最好的西欧亚祖先(并导致至少一位博客作者非常强烈地宣称东非人有西欧亚血统)与“Cardial”文化相同),ADMIXTURE 分析发现,这种血统最好解释为“西南亚”成分,在阿拉伯人中也很常见。
黎凡特新石器时代确实有这种和解,因为它与为欧洲人做出贡献的安纳托利亚新石器时代最接近。从而解释了正式的统计数据。
但与此同时,黎凡特新石器时代与 WHG 欧洲狩猎采集者的相关性明显低于安纳托利亚新石器时代,并且与 WHG 的相关程度可能与阿拉伯人群更相似。在模型中,论文发现安纳托利亚新石器时代的模型可建模约 40% WHG 和 60% 新石器时代(尽管我不确定他们的 33% 黎凡特和 33% 伊朗新石器时代模型是否真的有效)。
因此,这解释了为什么 ADMIXTURE 更喜欢用西南亚成分来模拟东非人,这些成分将他们与阿拉伯人联系起来(在欧洲地中海农民中发现缺乏欧洲 HG 血统/相关性)。
根据论文,Est 对 SNP 的数量和样本大小更敏感,达到某个点,但 Lazaridis 2016 集有足够的 SNP,并且对于许多古代人群来说,有足够的个体可以超过该点。有趣的是,Fst 似乎也很敏感,比较不同矩阵的结果?
无论如何,东部中位数似乎更符合正式统计数据,例如显示美洲原住民比约鲁巴人更接近欧洲人,并且密切相关但孤立的人群之间的距离没有扩大。
我想我对 ANE 传播的思考方式,以及将其地理覆盖范围置于上下文中,将被视为 ENA 分支的北部对应物,而不是与更广泛的旧石器时代晚期欧洲分支进行比较(基本上是 Fu 等人中的所有样本) 2016),WHG 就来自于此。
分裂的非洲东北部一侧正在从非洲向东扩展,而那个方向上有更多的无人居住的地区。因此,就像 ENA 分支一样,其后代覆盖了非常广阔的范围。也许这对我们来说似乎更令人难以置信,因为我们直到最近才知道这个组织的存在,而我们对 ENA 分支的范围已经相当熟悉了一段时间。
在我看来,ENA 祖先的主干很可能来自南方,穿过印度到达东亚,与此相比,ANE 今天在该范围内的生存边缘要大得多。我猜想这是因为北方人口遭受了更多的人口崩溃,然后被来自 ENA 分支的人口取代,这些人口适应了东亚大陆气候的凉爽气候(也许还有 ANE 变体的渗入)。
至于为什么 ANE 的存活率似乎通过 y 染色体系最强,也许我可以初步猜测,这是因为随着人口迁移到气候凉爽的地区,狩猎相对于采集策略变得越来越重要。来自 ANE 文化的雄性在文化/生物学上更适应在凉爽气候下狩猎,因此他们相关的 y-dna 具有更好的生存能力?
这非常有趣。从菲律宾和马来西亚、刚果、斯里兰卡、东非(“Dorobo”与尼罗河和库希特人口)和南非的狩猎采集者和农牧民之间的现代互动来看,人们可能会想到性别偏见的基因流动和文化影响,例如语言,只发生在一个方向,从农牧民到HGs,这总是使欧洲和西伯利亚的aDNA恢复情况,其中男性偏向的基因流动和文化主导地位似乎至少是在另一个方向有时,在社会上很难理解,因为没有什么可比较的。
由于您提到的原因,关于狩猎设施对两极的重要性日益增加的想法可以解释很多。此外,概括而言,在小规模社会中,肉类的交换和分享赋予了男性猎人巨大的地位。可能还指出了这样一个事实,即现代关于HG及其相互作用的知识偏向热带地区,这可能使我们对人类史前史的社会理解偏向于“黎明时的性”类型的平等主义场景,其中有大量的离婚和儿童独立,因为例如,远离爱斯基摩人和吉利亚克人等严格的家庭分化,这实际上可能更接近我们大多数欧亚人祖先的社会背景。
事实上,我现在不太愿意将基础欧亚人与单倍群 E 在非洲的传播联系起来,因为本文中的 D 统计数据显示撒哈拉以南非洲人(莫塔/约鲁巴/姆布蒂)与纳图夫人并不接近——后者是一半基础——甚至比 EHG 还要重要。无论如何,这都是一个奇怪的结果。
很棒的帖子!!!
--------------------------------------
WHG+EHG+EEF+CAF+异国情调(东亚/北亚、南亚、非洲)=现代中东和欧洲
这是正确的,但重要的是要记住 4 个古代西欧亚人群体中的每一个群体都存在差异。所有现代和古代的西欧亚人都没有来自这些大家族中完全相同的成员的祖先。这些大家庭中每个成员的具体关系尚不清楚。
例如,格鲁吉亚人不适合扎格罗斯新石器时代(CAF)+其他人。他们显然有很多血统,特别是来自高加索狩猎采集者,而不仅仅是来自扎格罗斯农民。此外,现代高加索人、安纳托利亚人和欧洲人显然有“EEF”血统,特别是来自安纳托利亚(或附近)而不是黎凡特。
中国有没有古代DNA?我只能找到这个:
http://bmcevolbiol.biomedcentral.com/articles/10.1186/1471-2148-13-216
这很有趣(单倍型N!),但每个人都想看的是仰韶对龙山,夏对夷,商朝到底是谁,他们从哪里得到战车,等等。
拉齐布
你知道有人会很快写一本关于这些发展的好书,如果不是很好的话,可以理解(正如智商低于 120 的群众可以理解的那样)? 如果是这样,你认为他们什么时候会发布它? 在没有某种可消化的记录的情况下,试图跟踪所有这些会引起头痛。
卡尔
我的观点是,多个语言群体的存在并不意味着东亚有多个人口从事农业,因为这些语言群体太浅,无法达到最早新石器时代所需的时间深度。 遗传证据——“东亚新石器时代星团”异常同质——似乎也与这一点相悖。
同样,巴布亚新几内亚的语言多样性很高,但从遗传学角度来看,“多种新石器时代”并没有得到保证。
语言多样性的存在与受地理因素影响而不受文化进化和高群体灭绝率影响的地区密切相关,例如高加索、喜马拉雅山脉南部和阿萨姆邦、东南亚高地、美国西北太平洋地区、亚马逊等。 我认为,在成熟的畜牧业和社会复杂性出现之前,欧亚大陆西部最早时期的语言多样性是可以预料到的。 事实上,我们的零假设应该是为什么中东在语言上不像新几内亚那样多样化,这将使我们得出这样的结论:自新石器时代后中东早期以来,群体竞争和高替代率就一直在运作,如 aDNA 中所示,与巴布亚不同。
我认为这篇论文表明,当今的遗传同质性不应导致人们推测东亚的新石器时代起源可能不是来自几个遗传上不同的群体。 无论如何,虽然东亚总体上相当同质,但汉藏族、南亚语族和南岛语族在基因上却足够独特,即使是简单的混合也能很好地在语言上将这些群体分开。 也许这种差异只是由于奥美拉尼西亚人基因渗入的比例不同或人口形成时期相当长的相对生殖隔离时期所致,但他们中的每一个相对于其他人确实仍然有一些独特之处。
回复:@Razib Khan
哦,顺便说一句,我的错......我最初误读了我发送的一些树混合。
在另一组树中,ryukendo 询问 Onge 分裂后,东亚人(但不是获得特定 ANE 的美洲原住民)和欧洲 HG/ANE 分支之间存在 16% 的优势。 我不会说树木排除了东亚人的 ASE+ANE 模型,但 Onge 或 ANE 本身可能有一段复杂的历史……"
是的,我想我们最终需要更多的数据和古代基因组(比如来自南亚和东亚的基因组)。 当我们拥有这些样本时,看看会发生什么应该很有趣。
回复:@PQ
将 ANE 输入到所有 ENA 祖先的模型涉及 WHG 和 ANE,作为来自以伊朗_N 为根基的更大基底混合组的最终提示; 此外,ANE 与 WHG 的边缘共享最多 3% 的漂移,即两者之间根本没有太大区别。 边缘很可能涉及 WHG 和 ANE 被拉近 ENA,因为需要考虑它们的非 Basal 与其他非 Basal 的接近程度。
不排除 ANE 和 ENA 有着复杂的历史; Chad 和我去检查了一些统计数据,其中 WHG-ANE 分支与 ENA 大致对称,但 MA-1 相对接近巴布亚人和 Onge,EHG 与东亚人狭义定义,一些 WHG 与东亚人狭义定义,SHG 不接近去任何; 但是 Treemix 中树的一个部分上的边缘可以是对另一部分上的现象的补偿性调整。
我的观点是,多种语言群体的存在并不意味着东亚有多种人口从事农业,因为这些语言群体太浅,无法达到最早新石器时代所需的时间深度。遗传证据——“东亚新石器时代群”异常同质——似乎也与这一点相悖。
同样,巴布亚新几内亚的语言多样性很高,但从遗传学角度来看,“多种新石器时代”并没有得到保证。
语言多样性的存在与受地理因素影响而不受文化进化和高群体灭绝率影响的地区密切相关,例如高加索、喜马拉雅山脉南部和阿萨姆邦、东南亚高地、美国西北太平洋地区、亚马逊等。我认为,在成熟的畜牧业和社会复杂性出现之前,欧亚大陆西部最早时期的语言多样性是可以预料到的。事实上,我们的零假设应该是为什么中东在语言上不像新几内亚那样多样化,这将使我们得出这样的结论:自新石器时代后中东早期以来,群体竞争和高替代率就一直在运作,如 aDNA 中所示,与巴布亚不同。
回复:@Karl Zimmerman
我认为这篇论文表明,当今的遗传同质性不应导致人们推测东亚的新石器时代起源可能不是来自几个遗传上不同的群体。 无论如何,虽然东亚总体上相当同质,但汉藏族、南亚语族和南岛语族在基因上却足够独特,即使是简单的混合也能很好地在语言上将这些群体分开。 也许这种差异只是由于奥美拉尼西亚人基因渗入的比例不同或人口形成时期相当长的相对生殖隔离时期所致,但他们中的每一个相对于其他人确实仍然有一些独特之处。
也许这种差异只是归因于奥美拉尼西亚人渗入的比例不同
不是全部。
另外,在东亚,Y chr 看起来像 J 或 E 一样爆裂,而不是 R1。我想我会同意附近有多个农民的想法。
回答古e。亚洲DNA。 q fu 正在努力。
我以为你支持最近的 OOA,Razib?如果现代人类迁徙了 OOA 50-60 kya,然后不久又迁徙回来,带来了 E 和 DE 的基础分支,那么他们必须与留在非洲的祖先非常相似(并且没有尼安德特人)。
我有点困惑发生了什么事。
我认为 OOA 可能早于 50-60 顺便说一句,尽管当时确实发生了欧亚大陆西南角的扩张。
现在对我来说可能并不疯狂的一个模型是,有一段时间存在三种流行音乐,SS非洲人,派生非非洲人,以及基础非“非洲人”,后者可能已经深入非洲,谁知道呢?这会让E在末次盛宴之后混合之前就地多样化吗?
但老实说,我不知道。
事实上,我现在不太愿意将基础欧亚人与单倍群 E 在非洲的传播联系起来,因为本文中的 D 统计数据显示撒哈拉以南非洲人(莫塔/约鲁巴/姆布蒂)与纳图夫人并不接近——后者是一半基础——甚至比 EHG 还要重要。无论如何,这都是一个奇怪的结果。
它必须非常男性化。显然,这个论点是双向的: 为什么欧亚大陆的 E 持有者与拥有 E 的非洲党卫军不更接近?
回复:@Razib Khan
我有点困惑发生了什么事。
我认为 OOA 可能早于 50-60 顺便说一句,尽管当时确实发生了欧亚大陆西南角的扩张。
现在对我来说可能并不疯狂的一个模型是,有一段时间存在三种流行音乐,SS非洲人,派生非非洲人,以及基础非“非洲人”,后者可能已经深入非洲,谁知道呢?这会让E在末次盛宴之后混合之前就地多样化吗?
但老实说,我不知道。
事实上,我现在不太愿意将基础欧亚人与单倍群 E 在非洲的传播联系起来,因为本文中的 D 统计数据显示撒哈拉以南非洲人(莫塔/约鲁巴/姆布蒂)与纳图夫人并不接近——后者是一半基础——甚至比 EHG 还要重要。无论如何,这都是一个奇怪的结果。
它必须非常男性化。显然,这个论点是双向的: 为什么欧亚大陆的 E 持有者与拥有 E 的非洲党卫军不更接近?
在这里尽量不要太过分,因为正式的统计数据没有显示纳图夫人中有任何非洲混血(根据对论文中未解决的样本的原始数据的进一步分析,这些人似乎都是 Y-DNA E1b1b)。
但在 Eurogenes 全球 PCA 中,Natufians 具有 SSA 拉力,与所有其他古代样本不同。在 Eurogenes TreeMix 中,它们有一些非常基本的迁移边缘,看起来像非洲。这与高度基础的欧亚裔伊朗人完全不同。他们的 Y-DNA 都是 E1b1b。即使没有混合东非参考资料,ADMIXTURE 也显示了其中的非洲成分。
一些东西告诉我,来自旧石器时代北非的古代 DNA 将揭示考古学家关于非洲对纳图夫人的影响的说法是正确的,只是他们并不完全像 SSA。然后,该元素将与真正的基底欧亚元素分开,从现有样品来看,基底欧亚元素似乎在中石器时代的伊朗达到顶峰。
然后,该元素将与真正的基底欧亚元素分开,从现有样品来看,基底欧亚元素似乎在中石器时代的伊朗达到顶峰。
但那里的 BEu 间隔很高。所以不确定它是否真的更高......
回复:@Shaikorth
我认为这篇论文表明,当今的遗传同质性不应导致人们推测东亚的新石器时代起源可能不是来自几个遗传上不同的群体。 无论如何,虽然东亚总体上相当同质,但汉藏族、南亚语族和南岛语族在基因上却足够独特,即使是简单的混合也能很好地在语言上将这些群体分开。 也许这种差异只是由于奥美拉尼西亚人基因渗入的比例不同或人口形成时期相当长的相对生殖隔离时期所致,但他们中的每一个相对于其他人确实仍然有一些独特之处。
回复:@Razib Khan
也许这种差异只是归因于奥美拉尼西亚人渗入的比例不同
不是全部。
另外,在东亚,Y chr 看起来像 J 或 E 一样爆裂,而不是 R1。我想我会同意附近有多个农民的想法。
回答古e。亚洲DNA。 q fu 正在努力。
但在 Eurogenes 全球 PCA 中,Natufians 具有 SSA 拉力,与所有其他古代样本不同。在 Eurogenes TreeMix 中,它们有一些非常基本的迁移边缘,看起来像非洲。这与高度基础的欧亚裔伊朗人完全不同。他们的 Y-DNA 都是 E1b1b。即使没有混合东非参考资料,ADMIXTURE 也显示了其中的非洲成分。
一些东西告诉我,来自旧石器时代北非的古代 DNA 将揭示考古学家关于非洲对纳图夫人的影响的说法是正确的,只是他们并不完全像 SSA。然后,该元素将与真正的基底欧亚元素分开,从现有样品来看,基底欧亚元素似乎在中石器时代的伊朗达到顶峰。
回复:@Razib Khan
然后,该元素将与真正的基底欧亚元素分开,从现有样品来看,基底欧亚元素似乎在中石器时代的伊朗达到顶峰。
但那里的 BEu 间隔很高。所以不确定它是否真的更高......
考虑到间隔,伊朗_N 的下层尼安德特人可能是一个指标。
回复:东亚新石器时代,ANE+ASE 的拟合似乎同样好,并且对于许多不同的群体来说比例相似,可能是他们祖先何时分裂的一个指标。
Yi、She、Kinh、Dai 和 Atayal 对于 Onge+MA-1 拟合具有相似的 P 值,并且每个人都获得 9-10% ANE。
然后,该元素将与真正的基底欧亚元素分开,从现有样品来看,基底欧亚元素似乎在中石器时代的伊朗达到顶峰。
但那里的 BEu 间隔很高。所以不确定它是否真的更高......
回复:@Shaikorth
考虑到间隔,伊朗_N 的下层尼安德特人可能是一个指标。
回复:东亚新石器时代,ANE+ASE 的拟合似乎同样好,并且对于许多不同的群体来说比例相似,可能是他们祖先何时分裂的一个指标。
Yi、She、Kinh、Dai 和 Atayal 对于 Onge+MA-1 拟合具有相似的 P 值,并且每个人都获得 9-10% ANE。
@拉齐布
如果我们在 K 值非常低的情况下观察 ADMIXTURE,我们会在欧亚大陆西部看到一个在印度河达到峰值的成分,另一个在地中海和黎凡特,另一个在波罗的海和伏尔加河地区;前两个组成部分与新石器时代基因组的纳图夫-安纳托利亚和甘吉达雷复合体非常匹配,后一个组成部分与北欧和西伯利亚狩猎采集生活方式的不成比例的生存相匹配。然后在欧亚大陆东部,我们在翁格和原住民中达到顶峰——这两个民族都是东南亚的狩猎采集者;另一个是在乙支/鄂伦春和恩加纳桑,再次是狩猎采集者(《阿穆尔》即将发表的论文可能会对东亚常染色体的这一元素有所启发),另一个涵盖了从日本到摩鹿加群岛之间的所有农牧民。这可能表明,即使存在多个新石器时代中心——我同意你的观点,不同新石器时代文化之间的Y染色体差异就是这一点的证据——它们在常染色体上的差异可能仍然比新石器时代基因组要小得多位于欧亚大陆西部。
东亚高K成分确实具有个体性,这表明过去几千年的人口流动;奇怪的是,这在 Y-DNA 记录中并不匹配,其中最成功的 O 分支的星状扩张可以追溯到新石器时代而不是更晚的时期。话又说回来,也许 J 仅存在于最初的新石器时代基因组中,这可能会让我们重新评估“类 J”扩展的作用。
分裂的非洲东北部一侧正在从非洲向东扩展,而那个方向上有更多的无人居住的地区。因此,就像 ENA 分支一样,其后代覆盖了非常广阔的范围。也许这对我们来说似乎更令人难以置信,因为我们直到最近才知道这个组织的存在,而我们对 ENA 分支的范围已经相当熟悉了一段时间。
在我看来,ENA 祖先的主干很可能来自南方,穿过印度到达东亚,与此相比,ANE 今天在该范围内的生存边缘要大得多。我猜想这是因为北方人口遭受了更多的人口崩溃,然后被来自 ENA 分支的人口取代,这些人口适应了东亚大陆气候的凉爽气候(也许还有 ANE 变体的渗入)。
至于为什么 ANE 的存活率似乎通过 y 染色体系最强,也许我可以初步猜测,这是因为随着人口迁移到气候凉爽的地区,狩猎相对于采集策略变得越来越重要。来自 ANE 文化的雄性在文化/生物学上更适应在凉爽气候下狩猎,因此他们相关的 y-dna 具有更好的生存能力?
回复:@Ryukendo
这非常有趣。从菲律宾和马来西亚、刚果、斯里兰卡、东非(“Dorobo”与尼罗河和库希特人口)和南非的狩猎采集者和农牧民之间的现代互动来看,人们可能会想到性别偏见的基因流动和文化影响,例如语言,只发生在一个方向,从农牧民到HGs,这总是使欧洲和西伯利亚的aDNA恢复情况,其中男性偏向的基因流动和文化主导地位似乎至少是在另一个方向有时,在社会上很难理解,因为没有什么可比较的。
由于您提到的原因,关于狩猎设施对两极的重要性日益增加的想法可以解释很多。此外,概括而言,在小规模社会中,肉类的交换和分享赋予了男性猎人巨大的地位。可能还指出了这样一个事实,即现代关于HG及其相互作用的知识偏向热带地区,这可能使我们对人类史前史的社会理解偏向于“黎明时的性”类型的平等主义场景,其中有大量的离婚和儿童独立,因为例如,远离爱斯基摩人和吉利亚克人等严格的家庭分化,这实际上可能更接近我们大多数欧亚人祖先的社会背景。
这有点题外话,但再次谈到巴布亚人的话题时,我从来没有完全理解新几内亚的 Y DNA 单倍群。
新几内亚东部最常见的群体是 K2b1 的分支,特别是 M 和 S。K2b1 基本上只在 Sahul 群体中发现。但 K2b1 的姐妹群是 P,它似乎是古代北欧亚单倍群。下一组 K1 包含 N 和 O,分别来自北极和东亚。
考虑到 K2b1 的外群和姐妹群似乎都起源于北亚,似乎最简单的假设是 K2b1 起源于北亚某个地方,只是后来才向南迁移。也许所有这一切都发生在萨胡尔定居之前,K2b1 与 C 单倍群一起并最终统治了澳大利亚。但两次迁移似乎也是合理的。
P1 (QR) 的近亲是 P2,发现于菲律宾内格里托人和帝汶岛。它的姐妹是 MS,分布在印度尼西亚东部和萨胡尔。
接下来是K2a/NO,但NO本身只是K2a的一个分支。 Pre-NO 发现于印度南部(也由 Ust'-Ishim Man 和 Oase-1 携带)。此外,K2*(xNO, MS) 在印度尼西亚西部发现,它一定代表某种前 NO 或 K2 的第三个分支(据我所知,从未测试过上游 K2a 标记)。印度尼西亚和印度都有需要正确分类的 K*。
所以不,MS 不嵌套在北欧亚单倍群中,P1 嵌套在印度尼西亚单倍群中。
然而,只有 1 个突变连接 K2a 和 K2b,1 个突变连接 MS 和 P,2 个突变连接 P1 和 P2,因此不涉及很大的时间深度。它们可能只是旧石器时代晚期 F 和 K 最初扩张的一部分,由于明显的原因,东南亚保存下来的谱系比西伯利亚多。
14年前发生了一次部分替代,然后是新石器时代的另一次,最后北欧人几乎全部被印欧人大规模替代,只留下一小部分狩猎采集者躲藏在森林和不适合放牧或耕种的地区。让我们看看这些女性或一些坚持不懈的 HG 做出了哪些贡献。
头发/眼睛颜色是浅色皮肤的次要基因,似乎都集中在欧洲,头发/眼睛颜色基因起源于十多千年前的欧洲土著。其中许多基因最近一定在欧洲,尤其是北欧重新繁荣起来。欧洲也缺乏具有足够优势而在其他地方受到广泛青睐的东西(东亚人的 EDAR 变体对下巴形状有深远的影响)。
至少对我来说,选择对外观产生影响的操作看起来是可行的。头发/眼睛的颜色似乎在北欧受到青睐,从征服缺乏眼睛头发颜色的印欧人开始,他们到达并杀死了几乎所有被征服的男人和许多女人。由此产生的欧洲男性/被征服女性的混合人口迅速变白,可能是因为白色皮肤被选择作为一种引起护理的特征。
在我看来,对于拥有 Y 染色体的北方猎人来说,作为主要的家庭供养者(至少在冬天),就是一夫一妻制。在北方,男性会因为试图建立和养活另一个家庭而降低他们的生殖健康。北方农业将会很艰难,而且粮食也有有限的缺点。在热带地区,性欲有限并不是一个优势,因为第二任妻子通常可以通过采集或园艺农业来获得。北方的 HG 男子很容易适应那里的农业。我想征服后的印欧人有他们对女性的选择,但也有一个日益严重的问题,即无法养活多个家庭。有些人必须离开,我认为这就是白皮肤从发射台上消失的地方。
在我看来
只需引用彼得·弗罗斯特感兴趣的论文即可。他比你清楚,而你只是像你一样不断重复他的想法。
回复:@Sean
新几内亚东部最常见的群体是 K2b1 的分支,特别是 M 和 S。K2b1 基本上只在 Sahul 群体中发现。但 K2b1 的姐妹群是 P,它似乎是古代北欧亚单倍群。下一组 K1 包含 N 和 O,分别来自北极和东亚。
考虑到 K2b1 的外群和姐妹群似乎都起源于北亚,似乎最简单的假设是 K2b1 起源于北亚某个地方,只是后来才向南迁移。也许所有这一切都发生在萨胡尔定居之前,K2b1 与 C 单倍群一起并最终统治了澳大利亚。但两次迁移似乎也是合理的。
回复:@Megalophias
P1 (QR) 的近亲是 P2,发现于菲律宾内格里托人和帝汶岛。它的姐妹是 MS,分布在印度尼西亚东部和萨胡尔。
接下来是K2a/NO,但NO本身只是K2a的一个分支。 Pre-NO 发现于印度南部(也由 Ust'-Ishim Man 和 Oase-1 携带)。此外,K2*(xNO, MS) 在印度尼西亚西部发现,它一定代表某种前 NO 或 K2 的第三个分支(据我所知,从未测试过上游 K2a 标记)。印度尼西亚和印度都有需要正确分类的 K*。
所以不,MS 不嵌套在北欧亚单倍群中,P1 嵌套在印度尼西亚单倍群中。
然而,只有 1 个突变连接 K2a 和 K2b,1 个突变连接 MS 和 P,2 个突变连接 P1 和 P2,因此不涉及很大的时间深度。它们可能只是旧石器时代晚期 F 和 K 最初扩张的一部分,由于明显的原因,东南亚保存下来的谱系比西伯利亚多。
@龙剑道
西欧亚成分在较低 K 下的更多差异似乎在 ADMIXTURE 面板上表现得最好,该面板的样本数量与西欧亚大陆相当不平衡,但我认为仍然正确,并且在东欧亚大陆和西欧亚大陆之间显示出更平衡的成分。我想你会期望,如果沿着离开非洲的路线,你会得到更广泛的密切相关进化枝的地理分布(就像欧亚大陆西部到非洲,到东亚,到美洲),而不是在东部出现令人惊讶的强烈漂移亚洲(与美洲不同)以及一个进化枝内更多的气候表型适应(正如移入美洲的相对密切相关的进化枝的情况)。
尽管即使在东亚大陆内部,人口之间的第一基础差异似乎仍然相当强大。很难找到包含大量西欧亚与西欧亚比较以及东亚与东亚比较的第一张大表。然而有一个来自 http://www.cell.com/ajhg/fulltext/S0002-9297(11)00488-5 –“南亚人口结构的共同和独特组成部分以及正选择的全基因组信号”。有些距离,如戴汉-法国-撒丁岛 (0.008),或戴-日本 0.019,与法国-贝都因人、格鲁吉亚-撒丁岛或俄罗斯-撒丁岛相当。
这可能是因为那里的样本来自具有不寻常漂流历史的少数民族,或者因为区域人口之间存在一些强烈的差异,这些差异不会落入大集群,因为它与个体人口不同(IRC东亚人口通常似乎下降)与树形结构中相对较长的单个分支未混合)。
当然,您也可以与 Shaikorth 谈到的最有效措施进行交叉核对。
回复:关于 HG 和农牧民之间互动的评论,我更多地想到了新石器时代前的 ANE 被西伯利亚的东亚群体取代,这似乎是一个在原始美国人离开前往美洲时已经部分完成的过程。美洲,然后发生了更进一步的事情,但这是新石器时代的一个有趣的延伸。
南汉和日本之间的中位数在格拉诺的补充中是 0.365。与欧亚大陆西部的可比距离为法国-卡尔戈波尔俄罗斯 0.319、贝加莫-托斯卡纳 0.245 和贝加莫-奥卡迪亚 0.455。柬埔寨-蒙古> 0.6,与卡拉什-俾路支相当。
OTOH 甚至傣语-日本等位基因共享距离(应对应于 IBS)也比法国-托斯卡纳或法国-奥卡迪亚等位基因短得多(Behar 等人 2010 年。补充)。这可能是由于东亚人总体上纯合性较高所致。
我的主要观点不是东亚或西欧亚大陆有或多或少的成分,这受到数据集中代表的人口的影响,我明白这一点,更多地指的是这样一个事实,在 ADMIXTURE 中与纯粹的西欧亚人或纯粹的东亚人,在 K=3 时,两个成分被纯西欧亚群中的新石器时代捕获,但在 K=3 时,东亚群中的推定新石器时代群体仅捕获了一个成分。在 K=4 时,东亚的新石器时代群体分裂为一个占主导地位的阿美族和另一个在东北亚人中的群体,但这两个组成部分与西欧亚大陆不同,与第一标准的差异并不大,其中一些差异可能来自于亚欧大陆的类似 Onge 的祖先。阿米/东南亚成分。随着 K 的增加,所有“东亚”成分彼此之间的 fst 都较低,当被迫到树上时形成一个单一的进化枝,但西欧亚成分通常分为西地中海/西南亚洲成分和 Gedrosia 成分/高加索类似的,来自 fsts 和它们被迫安装在树上。
我不确定追溯到非常古老的分支的第一次殖民和运动的纯合性将如何影响 ADMIXTURE,有趣的问题。看起来较高的 Ks 确实会受到较小基因座组中次要等位基因频率最近漂移的支配,并且可以提供更多关于最近发生的事件的信息。
顺便说一句,感谢 fst 和 est convo,以及引出的有趣的数字,这可能是由不同的漂移历史/多样性和不同分支之间的混合引起的,例如,我预计柬埔寨人会与类似 Onge 的基质混合,也许是除法/减法风格分析在这里也能有所帮助吗?
回复:@Matt_
西欧亚成分在较低 K 下的更多差异似乎在 ADMIXTURE 面板上表现得最好,该面板的样本数量与西欧亚大陆相当不平衡,但我认为仍然正确,并且在东欧亚大陆和西欧亚大陆之间显示出更平衡的成分。我想你会期望,如果沿着离开非洲的路线,你会得到更广泛的密切相关进化枝的地理分布(就像欧亚大陆西部到非洲,到东亚,到美洲),而不是在东部出现令人惊讶的强烈漂移亚洲(与美洲不同)以及一个进化枝内更多的气候表型适应(正如移入美洲的相对密切相关的进化枝的情况)。
尽管即使在东亚大陆内部,人口之间的第一基础差异似乎仍然相当强大。很难找到包含大量西欧亚与西欧亚比较以及东亚与东亚比较的第一张大表。然而,有一篇来自http://www.cell.com/ajhg/fulltext/S0002-9297(11)00488-5 - “南亚人类人口结构的共享和独特组成部分以及正选择的全基因组信号” 。有些距离,如戴汉-法国-撒丁岛 (0.008),或戴-日本 0.019,与法国-贝都因人、格鲁吉亚-撒丁岛或俄罗斯-撒丁岛相当。
这可能是因为那里的样本来自具有不寻常漂流历史的少数民族,或者因为区域人口之间存在一些强烈的差异,这些差异不会落入大集群,因为它与个体人口不同(IRC东亚人口通常似乎下降)与树形结构中相对较长的单个分支未混合)。
当然,您也可以与 Shaikorth 谈到的最有效措施进行交叉核对。
回复:关于 HG 和农牧民之间互动的评论,我更多地想到了新石器时代前的 ANE 被西伯利亚的东亚群体取代,这似乎是一个在原始美国人离开前往美洲时已经部分完成的过程。美洲,然后发生了更进一步的事情,但这是新石器时代的一个有趣的延伸。
回复:@Shaikorth、@Ryukendo
南汉和日本之间的中位数在格拉诺的补充中是 0.365。与欧亚大陆西部的可比距离为法国-卡尔戈波尔俄罗斯 0.319、贝加莫-托斯卡纳 0.245 和贝加莫-奥卡迪亚 0.455。柬埔寨-蒙古> 0.6,与卡拉什-俾路支相当。
OTOH 甚至傣语-日本等位基因共享距离(应对应于 IBS)也比法国-托斯卡纳或法国-奥卡迪亚等位基因短得多(Behar 等人 2010 年。补充)。这可能是由于东亚人总体上纯合性较高所致。
头发/眼睛颜色是浅色皮肤的次要基因,似乎都集中在欧洲,头发/眼睛颜色基因起源于十多千年前的欧洲土著。其中许多基因最近一定在欧洲,尤其是北欧重新繁荣起来。欧洲也缺乏具有足够优势而在其他地方受到广泛青睐的东西(东亚人的 EDAR 变体对下巴形状有深远的影响)。
至少对我来说,选择对外观产生影响的操作看起来是可行的。头发/眼睛的颜色似乎在北欧受到青睐,从征服缺乏眼睛头发颜色的印欧人开始,他们到达并杀死了几乎所有被征服的男人和许多女人。由此产生的欧洲男性/被征服女性的混合人口迅速变白,可能是因为白色皮肤被选择作为一种引起护理的特征。
在我看来,对于拥有 Y 染色体的北方猎人来说,作为主要的家庭供养者(至少在冬天),就是一夫一妻制。在北方,男性会因为试图建立和养活另一个家庭而降低他们的生殖健康。北方农业将会很艰难,而且粮食也有有限的缺点。在热带地区,性欲有限并不是一个优势,因为第二任妻子通常可以通过采集或园艺农业来获得。北方的 HG 男子很容易适应那里的农业。我想征服后的印欧人有他们对女性的选择,但也有一个日益严重的问题,即无法养活多个家庭。有些人必须离开,我认为这就是白皮肤从发射台上消失的地方。
回复:@Razib Khan
在我看来
只需引用彼得·弗罗斯特感兴趣的论文即可。他比你清楚,而你只是像你一样不断重复他的想法。
正如彼得所说,人口连续性并不存在,欧洲人似乎出现了混乱和大规模替换。我理解彼得的意思是,欧洲人的头发和眼睛的颜色加上白色的皮肤只是因为他们主要是北欧平原冰河时代土著祖先的后裔。但 DNA 研究表明,尽管他们的头发和眼睛颜色现在非常常见,但他们并非如此。
浅色/多样的头发和眼睛的颜色一定是在印欧征服时期爆发的。我们知道印欧人奴役了被征服的妇女,她们往往会遭遇可怕的命运,例如在葬礼上一次被牺牲几个人。白皮肤的外观显然是在印欧征服时期流行起来的,这就是为什么我认为这与在一个无情的等级制、父权制印欧社会中寻求照顾和提供妻子有关,在这个社会中,妇女的地位因无休止的征服而被降低,并且纳妾被俘的女性。
在使用彼得关于冰河时代的原始想法的同时,我解释了欧洲人目前的蓝眼睛和金发的外观是性选择第二次到来的结果。全白皮肤有点不同,但在印欧人出现并施加一种非常特殊类型的破坏后突然出现。
回复:@Razib Khan
在我看来
只需引用彼得·弗罗斯特感兴趣的论文即可。他比你清楚,而你只是像你一样不断重复他的想法。
回复:@Sean
正如彼得所说,人口连续性并不存在,欧洲人似乎出现了混乱和大规模替代。我理解彼得的意思是,欧洲人的头发和眼睛的颜色加上白色的皮肤只是因为他们主要是北欧平原冰河时代土著祖先的后裔。但 DNA 研究表明,尽管他们的头发和眼睛颜色现在非常常见,但他们并非如此。
浅色/多样化的头发和眼睛颜色一定是在印欧征服时期爆发的。我们知道印欧人奴役了被征服的妇女,她们往往会遭遇可怕的命运,例如在葬礼上一次被牺牲几个人。白皮肤的外观显然是在印欧征服时期流行起来的,这就是为什么我认为这与在一个无情的等级制、父权制印欧社会中寻求照顾和提供妻子有关,在这个社会中,妇女的地位因无休止的征服而被降低,并且纳妾被俘的女性。
在使用彼得关于冰河时代的原始想法的同时,我解释了欧洲人目前的蓝眼睛和金发的外观是性选择第二次到来的结果。全白皮肤有点不同,但在印欧人出现并施加一种非常特殊类型的破坏后突然出现。
或许。你所说的大部分内容都是相当推测的,而且看起来即使在你描述的一些事情发生很久之后,即使在初始阶段(例如,参见不列颠群岛报纸)之后,对浅色色素沉着的选择仍在继续。最终你没有证据,所以很难具体说。
此外,你往往对遗传学含糊其辞,这是不幸的。你应该解决这个问题。
正如彼得所说,人口连续性并不存在,欧洲人似乎出现了混乱和大规模替换。我理解彼得的意思是,欧洲人的头发和眼睛的颜色加上白色的皮肤只是因为他们主要是北欧平原冰河时代土著祖先的后裔。但 DNA 研究表明,尽管他们的头发和眼睛颜色现在非常常见,但他们并非如此。
浅色/多样的头发和眼睛的颜色一定是在印欧征服时期爆发的。我们知道印欧人奴役了被征服的妇女,她们往往会遭遇可怕的命运,例如在葬礼上一次被牺牲几个人。白皮肤的外观显然是在印欧征服时期流行起来的,这就是为什么我认为这与在一个无情的等级制、父权制印欧社会中寻求照顾和提供妻子有关,在这个社会中,妇女的地位因无休止的征服而被降低,并且纳妾被俘的女性。
在使用彼得关于冰河时代的原始想法的同时,我解释了欧洲人目前的蓝眼睛和金发的外观是性选择第二次到来的结果。全白皮肤有点不同,但在印欧人出现并施加一种非常特殊类型的破坏后突然出现。
回复:@Razib Khan
或许。你所说的大部分内容都是相当推测的,而且看起来即使在你描述的一些事情发生很久之后,即使在初始阶段(例如,参见不列颠群岛报纸)之后,对浅色色素沉着的选择仍在继续。最终你没有证据,所以很难具体说。
此外,你往往对遗传学含糊其辞,这是不幸的。你应该解决这个问题。
西欧亚成分在较低 K 下的更多差异似乎在 ADMIXTURE 面板上表现得最好,该面板的样本数量与西欧亚大陆相当不平衡,但我认为仍然正确,并且在东欧亚大陆和西欧亚大陆之间显示出更平衡的成分。我想你会期望,如果沿着离开非洲的路线,你会得到更广泛的密切相关进化枝的地理分布(就像欧亚大陆西部到非洲,到东亚,到美洲),而不是在东部出现令人惊讶的强烈漂移亚洲(与美洲不同)以及一个进化枝内更多的气候表型适应(正如移入美洲的相对密切相关的进化枝的情况)。
尽管即使在东亚大陆内部,人口之间的第一基础差异似乎仍然相当强大。很难找到包含大量西欧亚与西欧亚比较以及东亚与东亚比较的第一张大表。然而,有一篇来自http://www.cell.com/ajhg/fulltext/S0002-9297(11)00488-5 - “南亚人类人口结构的共享和独特组成部分以及正选择的全基因组信号” 。有些距离,如戴汉-法国-撒丁岛 (0.008),或戴-日本 0.019,与法国-贝都因人、格鲁吉亚-撒丁岛或俄罗斯-撒丁岛相当。
这可能是因为那里的样本来自具有不寻常漂流历史的少数民族,或者因为区域人口之间存在一些强烈的差异,这些差异不会落入大集群,因为它与个体人口不同(IRC东亚人口通常似乎下降)与树形结构中相对较长的单个分支未混合)。
当然,您也可以与 Shaikorth 谈到的最有效措施进行交叉核对。
回复:关于 HG 和农牧民之间互动的评论,我更多地想到了新石器时代前的 ANE 被西伯利亚的东亚群体取代,这似乎是一个在原始美国人离开前往美洲时已经部分完成的过程。美洲,然后发生了更进一步的事情,但这是新石器时代的一个有趣的延伸。
回复:@Shaikorth、@Ryukendo
我的主要观点不是东亚或西欧亚大陆有或多或少的成分,这受到数据集中代表的人口的影响,我明白这一点,更多地指的是这样一个事实,在 ADMIXTURE 中与纯粹的西欧亚人或纯粹的东亚人,在 K=3 时,两个成分被纯西欧亚群中的新石器时代捕获,但在 K=3 时,东亚群中的推定新石器时代群体仅捕获了一个成分。在 K=4 时,东亚的新石器时代群体分裂为一个占主导地位的阿美族和另一个在东北亚人中的群体,但这两个组成部分与西欧亚大陆不同,与第一标准的差异并不大,其中一些差异可能来自于亚欧大陆的类似 Onge 的祖先。阿米/东南亚成分。随着 K 的增加,所有“东亚”成分彼此之间的 fst 都较低,当被迫到树上时形成一个单一的进化枝,但西欧亚成分通常分为西地中海/西南亚洲成分和 Gedrosia 成分/高加索类似的,来自 fsts 和它们被迫安装在树上。
我不确定追溯到非常古老的分支的第一次殖民和运动的纯合性将如何影响 ADMIXTURE,有趣的问题。看起来较高的 Ks 确实会受到较小基因座组中次要等位基因频率最近漂移的支配,并且可以提供更多关于最近发生的事件的信息。
顺便说一句,感谢 fst 和 est convo,以及引出的有趣的数字,这可能是由不同的漂移历史/多样性和不同分支之间的混合引起的,例如,我预计柬埔寨人会与类似 Onge 的基质混合,也许是除法/减法风格分析在这里也能有所帮助吗?
啊,谢谢你的解释,我现在更明白你在说什么了。 IIUC:在任何给定的西欧亚人/东亚人群体的 K 下,东亚出现的与任何新石器时代考古扩张相关的成分较少,并且在与新石器时代考古扩张相关的成分之间存在更紧密的区分。出现。
我个人无法对此做出真正的判断,因为我不确定我是否亲自见过任何具有可比样本大小/地理广度的纯西欧亚人口和纯东亚人口进行比较,而且绝对没有与已发布的组件之间的 fsts(我很确定我对新石器时代考古学的了解较少),但我会接受您对此的评论。
回复:@PQ
我的主要观点不是东亚或西欧亚大陆有或多或少的成分,这受到数据集中代表的人口的影响,我明白这一点,更多地指的是这样一个事实,在 ADMIXTURE 中与纯粹的西欧亚人或纯粹的东亚人,在 K=3 时,两个成分被纯西欧亚群中的新石器时代捕获,但在 K=3 时,东亚群中的推定新石器时代群体仅捕获了一个成分。在 K=4 时,东亚的新石器时代群体分裂为一个占主导地位的阿美族和另一个在东北亚人中的群体,但这两个组成部分与西欧亚大陆不同,与第一标准的差异并不大,其中一些差异可能来自于亚欧大陆的类似 Onge 的祖先。阿米/东南亚成分。随着 K 的增加,所有“东亚”成分彼此之间的 fst 都较低,当被迫到树上时形成一个单一的进化枝,但西欧亚成分通常分为西地中海/西南亚洲成分和 Gedrosia 成分/高加索类似的,来自 fsts 和它们被迫安装在树上。
我不确定追溯到非常古老的分支的第一次殖民和运动的纯合性将如何影响 ADMIXTURE,有趣的问题。看起来较高的 Ks 确实会受到较小基因座组中次要等位基因频率最近漂移的支配,并且可以提供更多关于最近发生的事件的信息。
顺便说一句,感谢 fst 和 est convo,以及引出的有趣的数字,这可能是由不同的漂移历史/多样性和不同分支之间的混合引起的,例如,我预计柬埔寨人会与类似 Onge 的基质混合,也许是除法/减法风格分析在这里也能有所帮助吗?
回复:@Matt_
啊,谢谢你的解释,我现在更明白你在说什么了。 IIUC:在任何给定的西欧亚人/东亚人群体的 K 下,东亚出现的与任何新石器时代考古扩张相关的成分较少,并且在与新石器时代考古扩张相关的成分之间存在更紧密的区分。出现。
我个人无法对此做出真正的判断,因为我不确定我是否亲自见过任何具有可比样本大小/地理广度的纯西欧亚人口和纯东亚人口进行比较,而且绝对没有与已发布的组件之间的 fsts(我很确定我对新石器时代考古学的了解较少),但我会接受您对此的评论。
其中一些分析是由相当不起眼的爱好者完成的,通常是在很久以前。大多数其他人并不关注 HUGO PASNP 数据集,因为它与其他数据集的重叠度非常差。
以下是第欧根尼·阿耳忒弥斯 (Diogenes Artemis) 的一些组件之间的对比:
http://4.bp.blogspot.com/-I9bLmnKcUtY/Te0_m4cl14I/AAAAAAAAAKY/w1vg_6YUb_E/s1600/Screenshot-2.png
深蓝色、红色、浅绿色和深绿色分别是“南亚语系”、“东北亚语系”、“苗族语系”和“南岛语系”分量,它们之间的 fst 徘徊在 05 左右。这与他在非洲内部运行中的多个“班图”组件之间的情况大致相同。
来自菲律宾的博主:
http://1.bp.blogspot.com/-2of7N2PmkIw/UmIJT81VglI/AAAAAAAAAcI/KLmw0z5OT-I/s1600/SILANGAN11SL90007KU2GP.19.Fst.JPG
http://4.bp.blogspot.com/-8Fkc2gRdka4/Uk95yU-fKlI/AAAAAAAAAb4/FO1saQoum3E/s1600/image002.png
虽然西欧亚成分(其中 4 个成分)之间的差异约为 5,但 3 个东亚成分之间的差异仅为约 25,只有西伯利亚/北亚成分之间的差异约为 5。 还要注意西欧亚成分是如何分为地中海-西亚洲的区别的,我们现在对最终原因有了更好的了解,但东亚农业学家处于一个分支中:
http://2.bp.blogspot.com/-RHaCKCcUNCo/Uk95yRJcFUI/AAAAAAAAAbs/UPUrXasHkMI/s1600/image001.png
非常有趣的文章。谢谢。
我唯一的评论是,我希望看到更多此类内容,不一定只涉及遗传学或科学,而是与智力相关。在乌兹别克斯坦大学发表的帖子(以及随后的评论帖子)中占主导地位的政治咆哮通常是空洞和令人厌烦的。
啊,谢谢你的解释,我现在更明白你在说什么了。 IIUC:在任何给定的西欧亚人/东亚人群体的 K 下,东亚出现的与任何新石器时代考古扩张相关的成分较少,并且在与新石器时代考古扩张相关的成分之间存在更紧密的区分。出现。
我个人无法对此做出真正的判断,因为我不确定我是否亲自见过任何具有可比样本大小/地理广度的纯西欧亚人口和纯东亚人口进行比较,而且绝对没有与已发布的组件之间的 fsts(我很确定我对新石器时代考古学的了解较少),但我会接受您对此的评论。
回复:@PQ
其中一些分析是由相当不起眼的爱好者完成的,通常是在很久以前。 大多数其他人并不关注 HUGO PASNP 数据集,因为它与其他数据集的重叠度非常差。
以下是第欧根尼·阿耳忒弥斯 (Diogenes Artemis) 的一些组件之间的对比:
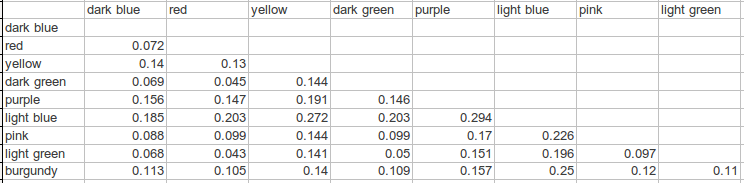
深蓝色、红色、浅绿色和深绿色分别是“南亚语系”、“东北亚语系”、“苗族语系”和“南岛语系”分量,它们之间的 fst 徘徊在 05 左右。 这与他在非洲内部运行中的多个“班图”组件之间的情况大致相同。
来自菲律宾的博主:
虽然西欧亚成分(其中 4 个成分)之间的差异约为 5,但 3 个东亚成分之间的差异仅为约 25,只有西伯利亚/北亚成分之间的差异约为 5。 还要注意西欧亚成分是如何分为地中海-西亚洲的区别的,我们现在对最终原因有了更好的了解,但东亚农业学家处于一个分支中:
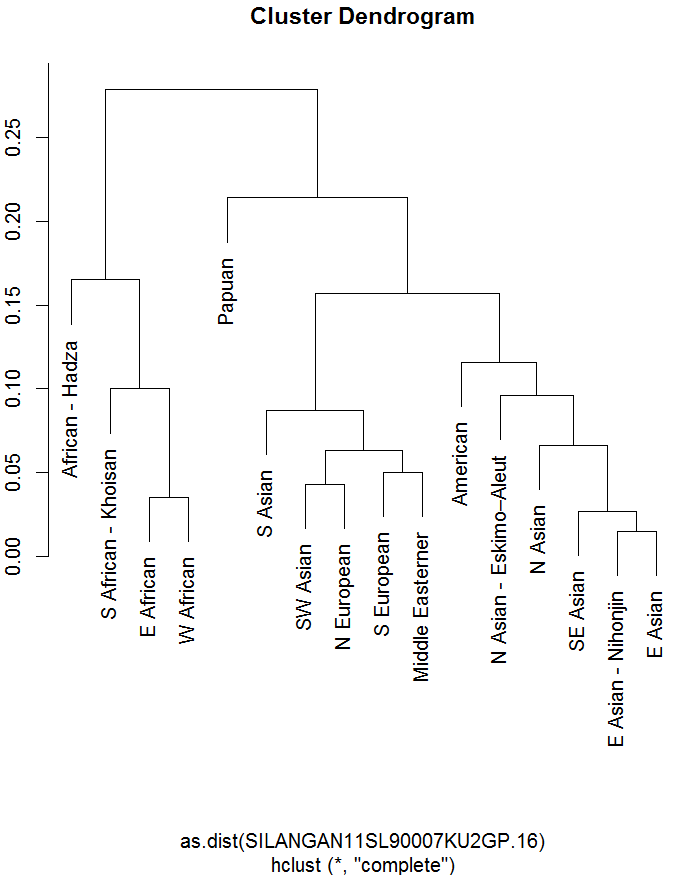
这些活动涵盖了从西伯利亚到巴布亚的人口,也包括其间的所有“黑人”,所以我认为代表性应该相当好。 菲律宾博主创建了基诺组件,即汉藏组件! 我想这是我唯一一次看到这样的事情。
顺便说一句,这是 RK,以不同网络的不同名称发帖,抱歉有多个绰号。