[zenphotopress相册= 249排序=订单编号= 4]
 首先,我想指出,我会不时为家人使用缩写(这适用于以后的帖子)。 我的父亲将是RF,我的母亲将是RM,我的兄弟姐妹将是RS,并用数字来表示哪个兄弟姐妹。 所以目前我们有RS1。 如您所见 完形 感觉我们就像一个家庭一样彼此很像。 我们大约有40%的亚洲人和60%的欧洲人。 碎片的程度表明我们不是最近的混合物。 否则,亚洲和欧洲碎片将聚集在一根或另一根铁线上。 有人建议我母亲的确表现出较少的支离破碎。 可能是这样的假设是,她的祖父据说来自中东的一个家庭,该家庭定居在南亚,先是在德里,然后定居在孟加拉东南部(具体来说是孟加拉国)。 诺哈里)。 由于根据23andMe的算法,他可能几乎没有亚洲血统,因此从他那里继承来的同系物绝大多数是欧洲人,只有一代人会进行重组。
首先,我想指出,我会不时为家人使用缩写(这适用于以后的帖子)。 我的父亲将是RF,我的母亲将是RM,我的兄弟姐妹将是RS,并用数字来表示哪个兄弟姐妹。 所以目前我们有RS1。 如您所见 完形 感觉我们就像一个家庭一样彼此很像。 我们大约有40%的亚洲人和60%的欧洲人。 碎片的程度表明我们不是最近的混合物。 否则,亚洲和欧洲碎片将聚集在一根或另一根铁线上。 有人建议我母亲的确表现出较少的支离破碎。 可能是这样的假设是,她的祖父据说来自中东的一个家庭,该家庭定居在南亚,先是在德里,然后定居在孟加拉东南部(具体来说是孟加拉国)。 诺哈里)。 由于根据23andMe的算法,他可能几乎没有亚洲血统,因此从他那里继承来的同系物绝大多数是欧洲人,只有一代人会进行重组。
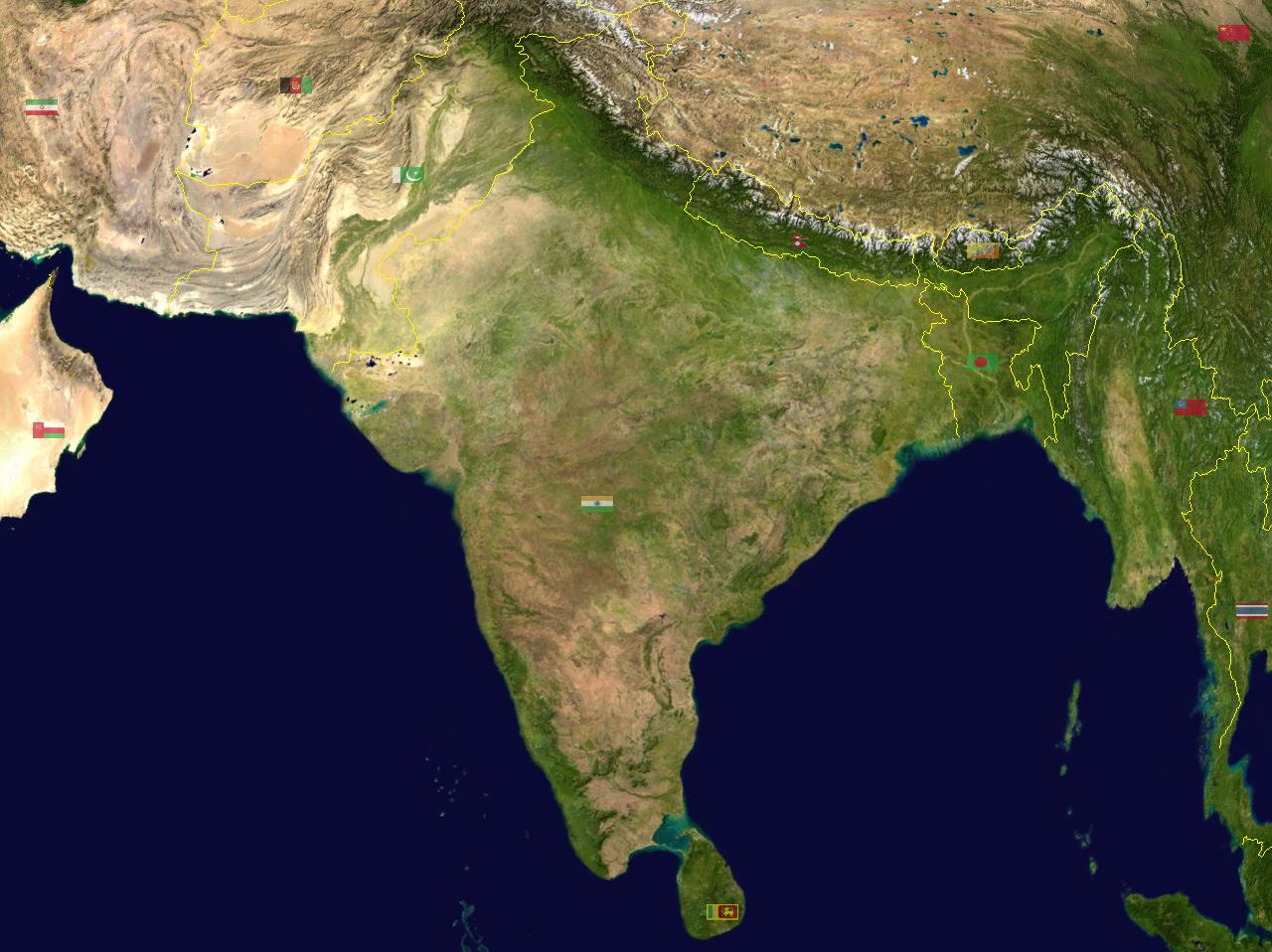
要评估各种假设的合理性以解释结果的模式,您需要所有非基因组信息。 上面是英属印度的地图。 我已经指出了我的家人来自孟加拉的地区。 在我的曾祖父母中,八分之七出生于 库米拉 (实际上,它比目前的孟加拉国行政区划更向达卡东南延伸)。 一位祖父母在科米利亚东南部的诺阿卡利(Noakhali)出生。 1位曾曾祖父母中有4位出生在距小镇8英里内 坚德布尔 (RF的祖父母)。 3个曾祖父母中有4个是在距该村庄5英里内出生的 霍姆纳 (RM的祖父母)。 乌鸦飞过时,这两个位置相距约30英里,不过在更早的时代,这两个位置之间的运输是靠水进行的。 梅格纳 河流,到恒河到达孟加拉国时,实际上比恒河的水量要大得多)。 这个地区以西部为边界 帕德玛河,在Chandpur的宽度缩小到大约2英里(平均深度约1,000英尺)。 东边是印度的 特里普拉邦。 这是地图上定义的相对较疏松的边界,不受地理因素强加。 您可以看到,在某些地区,实际上东部的Bangaldesh-印度边界 二等分稻田.

Tripuri儿童
如今,由于印度教徒从20世纪的东孟加拉邦(后来的东巴基斯坦,现在的孟加拉国)大规模迁移,特里普拉邦已成为孟加拉的多数民族。 但是它的土著人民是 特里普利,其母语显然是部落 藏缅人和物理类型指向它们与北部和东部人口的联系。 同时,约90%的Tripuri是印度教徒,在南亚伊斯兰统治时期, 特里普拉王朝 自命为印度教文明的捍卫者(就像阿萨姆邦的藏缅人阿洪斯所做的那样)。 因此,从语言和基因上讲,特里普拉邦的原住民与恒河平原的印度-雅利安民族形成鲜明的对比,孟加拉人是阿萨姆人最东端的代表。 但是,它们也早已成为南亚文化景观的一部分,因此不能再被视为纯粹的侵入性(它们的口述历史表明它们已经到来了。 before 穆斯林)。
最后,关于我的8位曾祖父母的详细背景,其中2位属于可汗阶层。 1个来自印度教家庭 塔库尔 最近recently依伊斯兰教的人。 另一个是姓氏 萨卡。 至少有一部分来自中东移植家庭中的1名。 剩下的4个曾祖父母是孟加拉穆斯林,除了我父母所知道的背景信息外,没有其他特别的背景信息。
我都给你了 因为遗传变异是 非常 以地域为条件 和 文化参数。 在前现代时期,水障似乎在文化和遗传上将人们分隔开来特别有效(尽管具有讽刺意味的是,水还是任何大宗贸易的先决条件)。 语言也是差异的另一个主要参数。 最后是宗教。 在上一节中,如果300年前我这一代的大多数祖先是印度教徒,我不会感到惊讶。 这显然有一些流动性。 我提供了出生地半径的数据,因为我们知道 欧洲成绩 这 甚至村庄也表现出遗传聚类。 这在我的家庭中得到了缓解,因为就社区而言,父亲的祖父母背景各异,而母亲的祖父母则来自不同的地区,从生物学上说,在很大程度上是不同的族裔。
 当我最初看到自己是40%的亚洲人时,我对这个高比例感到惊讶(记住,南亚的平均水平约为25%的亚洲人),但是有两种par昧的解释:a)我有很多ASI, b)我的祖先看上去并不像南亚那样,但确实来自东亚。 为了确定是否是前者,我在23andMe上开始主动与众多南亚人共享基因。 经过数十个人的了解, 我不在正常的变异间隔内。 我比来自南印度或斯里兰卡的人更亚洲。 此外,在HGDP数据集中,即使这些个体在基因上也往往最接近中南/亚洲人。 我最接近东亚人。 此外,在投影到中南/亚洲人的二维PCA上,我肯定不在所有其他南亚人的群体中。 最终,我确实找到了一个打破35%的亚洲魔力壁垒的人……那个人是孟加拉国人,占38%。 而且,像我一样,他在基本的“全球相似性”比赛中与东亚人更加接近。 他还携带了一个 Y染色体谱系 在南亚很少见,在苗族中很常见。 终于,当狄尼克斯(Dienekes)开始他的 Dodecad祖先项目 很明显,我的祖先中约有15%的元素与 不能 南亚,但东亚。 如果除去这一部分,那么我将是大约70%的欧洲人和30%的亚洲人,对于在该次大陆东部或南部有祖先的人来说,这绝对在正常范围内。
当我最初看到自己是40%的亚洲人时,我对这个高比例感到惊讶(记住,南亚的平均水平约为25%的亚洲人),但是有两种par昧的解释:a)我有很多ASI, b)我的祖先看上去并不像南亚那样,但确实来自东亚。 为了确定是否是前者,我在23andMe上开始主动与众多南亚人共享基因。 经过数十个人的了解, 我不在正常的变异间隔内。 我比来自南印度或斯里兰卡的人更亚洲。 此外,在HGDP数据集中,即使这些个体在基因上也往往最接近中南/亚洲人。 我最接近东亚人。 此外,在投影到中南/亚洲人的二维PCA上,我肯定不在所有其他南亚人的群体中。 最终,我确实找到了一个打破35%的亚洲魔力壁垒的人……那个人是孟加拉国人,占38%。 而且,像我一样,他在基本的“全球相似性”比赛中与东亚人更加接近。 他还携带了一个 Y染色体谱系 在南亚很少见,在苗族中很常见。 终于,当狄尼克斯(Dienekes)开始他的 Dodecad祖先项目 很明显,我的祖先中约有15%的元素与 不能 南亚,但东亚。 如果除去这一部分,那么我将是大约70%的欧洲人和30%的亚洲人,对于在该次大陆东部或南部有祖先的人来说,这绝对在正常范围内。
 如果您已经读到这一点, 您可能想知道我的父亲是38%的亚洲人,母亲是40%的亚洲人,而我是43%的亚洲人。 毕竟,我不应该成为 两者之间? 实际上,在PCA散布图上,我(和我的兄弟姐妹一样)正好在父母之间(由于标志太大,所以看不到后代)。 那么为什么会有所不同呢? 首先,请记住,PCA将您投影到二维轴上,其中x和y代表数据集中方差的两个最大部分。 换句话说,它正在淘汰 子集 基因变异的确在群体差异之间脱颖而出。 这样一来,作为第一代欧亚人的人就可以在这个情节上与父母相距甚远,但在总基因组方面仍然表现出很大的身份认同。 二维图无法捕获很多变化(例如,家族谱系的私人变体)。 祖先绘画的估计是不同的。 他们正在寻找 全 基因组,并评估每个区域在三个参考人群之间的遗传亲和力。 因此,请重复一遍,您拥有超过50个参考群体与3个群体,并且相对于整个基因组,您占总遗传变异的一小部分。 两种方法都报告了真实有效的结果,但是有些不同。
如果您已经读到这一点, 您可能想知道我的父亲是38%的亚洲人,母亲是40%的亚洲人,而我是43%的亚洲人。 毕竟,我不应该成为 两者之间? 实际上,在PCA散布图上,我(和我的兄弟姐妹一样)正好在父母之间(由于标志太大,所以看不到后代)。 那么为什么会有所不同呢? 首先,请记住,PCA将您投影到二维轴上,其中x和y代表数据集中方差的两个最大部分。 换句话说,它正在淘汰 子集 基因变异的确在群体差异之间脱颖而出。 这样一来,作为第一代欧亚人的人就可以在这个情节上与父母相距甚远,但在总基因组方面仍然表现出很大的身份认同。 二维图无法捕获很多变化(例如,家族谱系的私人变体)。 祖先绘画的估计是不同的。 他们正在寻找 全 基因组,并评估每个区域在三个参考人群之间的遗传亲和力。 因此,请重复一遍,您拥有超过50个参考群体与3个群体,并且相对于整个基因组,您占总遗传变异的一小部分。 两种方法都报告了真实有效的结果,但是有些不同。
 因此,对于以上差异,我可以想到两种非常简单的方法论解释。 我在V2上,而我的父母和兄弟姐妹在V3上。 我知道这在其他度量方面有所不同。 此外,此算法中显然存在一些“杂音”,即使您考虑了参考人群的k杂性质,也会导致具有非洲或亚洲血统的人是不真实的。 但是,让我们以结果为准。 用祖先画 回想一下欧洲和亚洲的情况 矮胖 整个基因组? 我的两个父母都从父母那里获得了一半的基因组。 我自己的染色体是祖父母的染色体的镶嵌体。 基因组区域之间的某些原始链接由于其在同一链上的物理位置而被打破, 重组 在我祖父母的下游两代人中。 具体而言,减数分裂的两个实例会产生性细胞。 因此,我祖父母中存在的一些等位基因关联在我内部已经转化。 但是,即使没有重组,也很明显,一条同源染色体在欧洲或亚洲可能比总基因组平均值要多。 因为只有其中一个被传递给任何给定的后代,所以从同级到同级会有差异。 遗传学不是一个纯粹的融合过程。 这就是为什么我是43%的亚洲人,而RS1是40%的亚洲人的原因。 我们都从父母的基因中取样,并且在这个过程中会有差异(在染色体水平上,您从每个父母那里获得22个常染色体引物,每个引物都有两个结果)。
因此,对于以上差异,我可以想到两种非常简单的方法论解释。 我在V2上,而我的父母和兄弟姐妹在V3上。 我知道这在其他度量方面有所不同。 此外,此算法中显然存在一些“杂音”,即使您考虑了参考人群的k杂性质,也会导致具有非洲或亚洲血统的人是不真实的。 但是,让我们以结果为准。 用祖先画 回想一下欧洲和亚洲的情况 矮胖 整个基因组? 我的两个父母都从父母那里获得了一半的基因组。 我自己的染色体是祖父母的染色体的镶嵌体。 基因组区域之间的某些原始链接由于其在同一链上的物理位置而被打破, 重组 在我祖父母的下游两代人中。 具体而言,减数分裂的两个实例会产生性细胞。 因此,我祖父母中存在的一些等位基因关联在我内部已经转化。 但是,即使没有重组,也很明显,一条同源染色体在欧洲或亚洲可能比总基因组平均值要多。 因为只有其中一个被传递给任何给定的后代,所以从同级到同级会有差异。 遗传学不是一个纯粹的融合过程。 这就是为什么我是43%的亚洲人,而RS1是40%的亚洲人的原因。 我们都从父母的基因中取样,并且在这个过程中会有差异(在染色体水平上,您从每个父母那里获得22个常染色体引物,每个引物都有两个结果)。
一个有趣的含义是,多种族夫妇的孙子在主要种族群体的祖先数量上将表现出差异。 这就是为什么认为通婚会导致生物多样性被洗掉是谬论的原因之一。 而且诸如分类交配的过程甚至可能从最初混合的随机交配种群中提取出“纯”个体。
尽管如此, 我现在相信,由于孟加拉东部的N = 3,所以我在最近的东南亚血统中也不例外,但孟加拉东部是南亚和东南亚之间的基因频率谱系的一部分,因此在很大程度上东方血统。 Zack拥有我父母的数据,因此一旦结果从 行动计划 我相信他会在狄恩凯克斯身上发现与我相同的非南亚血统。 这里的上升趋势仍然很尖锐。 在推断祖先量子算法的比例时,孟加拉国的平均水平可能与缅甸人的平均水平只有10-20%的互换性。 (请记住,缅甸人也可能有一个较小的南亚部分)。 相比之下,孟加拉国的平均水平可能与比哈尔邦居民保持80-90%的互换性(我与之共享的23andMe中SNP总比较中最接近的匹配项是比哈尔邦,而不是其他两个孟加拉国)。 显然,这是 地理, 南北 缅甸的山脉将其与南亚隔离开来。 相反,有 开阔的平原 从孟加拉国北部到比哈尔邦。 在某种程度上,由于海上旅行的便利,缅甸与南亚半岛之间有着更多的文化亲和力和联系。 患病率 上座部佛教 在缅甸证明了下层社会的联合 伊洛瓦底江 斯里兰卡。
 回到孟加拉国。 就宗教人口统计而言,印度次大陆的一个方面是印度伊斯兰教的心脏地带,德里地区,从来没有穆斯林占多数。 相反,在西北和东北边缘(以及其他一些地区,例如喀拉拉邦北部),穆斯林占多数。 伊斯兰教在西北地区的盛行并不令人惊讶,因为该地区与伊斯兰教徒接壤。 达拉伊斯兰教 恰当的。 但是孟加拉呢? 在19世纪后期,英国人感到惊讶的是,在统一的孟加拉国(大致包括印度的西孟加拉邦和孟加拉国),穆斯林占多数。 由于出生率和转换率的差异(如上所述,第二部分包括我的家庭部分),今天活着的孟加拉人中有三分之二是穆斯林,而其余的是印度教徒。 孟加拉国估计是2%的穆斯林,而西孟加拉邦是3%的穆斯林。 即使在几代印度教徒移民之后的今天,孟加拉国内部的一种模式仍然是印度教徒在西方和北部的相对集中度(而且,孟加拉国的印度教徒往往是城市化的)。 孟加拉国“古兰经”带的“带扣”实际上是孟加拉东南边缘的Noakhali区。 我母亲的外公,来自 PIRS 最初定居在德里的穆斯林心脏地带的诺亚里(Noakhali)。 显然,据说在Noakhali甚至印度教徒都知道适当的伊斯兰教形式!**
回到孟加拉国。 就宗教人口统计而言,印度次大陆的一个方面是印度伊斯兰教的心脏地带,德里地区,从来没有穆斯林占多数。 相反,在西北和东北边缘(以及其他一些地区,例如喀拉拉邦北部),穆斯林占多数。 伊斯兰教在西北地区的盛行并不令人惊讶,因为该地区与伊斯兰教徒接壤。 达拉伊斯兰教 恰当的。 但是孟加拉呢? 在19世纪后期,英国人感到惊讶的是,在统一的孟加拉国(大致包括印度的西孟加拉邦和孟加拉国),穆斯林占多数。 由于出生率和转换率的差异(如上所述,第二部分包括我的家庭部分),今天活着的孟加拉人中有三分之二是穆斯林,而其余的是印度教徒。 孟加拉国估计是2%的穆斯林,而西孟加拉邦是3%的穆斯林。 即使在几代印度教徒移民之后的今天,孟加拉国内部的一种模式仍然是印度教徒在西方和北部的相对集中度(而且,孟加拉国的印度教徒往往是城市化的)。 孟加拉国“古兰经”带的“带扣”实际上是孟加拉东南边缘的Noakhali区。 我母亲的外公,来自 PIRS 最初定居在德里的穆斯林心脏地带的诺亚里(Noakhali)。 显然,据说在Noakhali甚至印度教徒都知道适当的伊斯兰教形式!**
这种模式的一种解释是,印度教精英的宗教影响力和权力随着距离西孟加拉邦(更接近核心地区)的距离的直接函数而下降。 阿亚瓦拉塔在伊斯兰教兴起之前,传统上一直是印度历代王朝的权力所在地。 此外,孟加拉国是大陆次大陆的最后一个区域,在佛教徒开花期,佛教社会蓬勃发展。 帕拉帝国 大约在公元1000年左右。因此,建议许多孟加拉穆斯林是直接从佛教而不是印度教转变而来的(直到今天,仍有少数少数民族的孟加拉佛教徒,他们的姓氏是“包拉”。这是与后裔的区别)现在讲孟加拉语但仍保留部落身份和Theravada佛教徒的藏缅人)。 另外,可能是在穆斯林到来之前,孟加拉东部地区的泛灵论者居多,就像欧洲殖民大国在亚洲更成功地在边缘化人群中传播其宗教一样(例如,印度东北部和东南亚的部落人民伊斯兰教徒),因此伊斯兰教在印度教种姓制度之外的人中进行购买。
这些模型对我来说具有说服力。 但是,我仍然感到怀疑的是,在孟加拉西部和东部,印度教机构的深度存在如此强烈的脱节。 毕竟,当伊斯兰教在南亚刚刚出现时,东方的Tripura国王就是印度教徒。 如果在核心的印度教文明中处于部落和边缘地位是对伊斯兰教易感的理由之一,那么奇怪的是, 恰恰是现代孟加拉国的许多部落人民 不能 穆斯林。 的确,在南亚东部接近印度裔群体的藏缅人口是印度教徒或佛教徒,而不是穆斯林(腹地的那些人并未融入任何南亚宗教,而是在上个世纪由西方传教士转变为基督教)。
相反,我发现模型支持 伊斯兰兴起与孟加拉国边境,1204-1760年 作为上述假设的补充,甚至替代,在很大程度上是合理的。 此外,它还具有使我到目前为止在这里介绍的遗传数据有意义的作用。 作者认为 孟加拉国东部的孟加拉东部地区,在13世纪穆斯林占领孟加拉之前,人口非常稀少。 在现代时期,印度的孟加拉西部地区往往遇到许多水道濒临灭绝的问题。 但是在一千年前,该地区在沉积方面更为活跃,而孟加拉东部是一个荒野。 几个世纪以来,大河向东转移,由于运输的改善,该地区得以开垦。 此外,穆斯林的到来也导致了新的土地清理和定居技术的传播。 粗略的模型是 就目前的人口密度而言,孟加拉东部实际上是一个相对较新的定居地区。 由于清算和定居活动是由穆斯林精英进行的,因此,定居在这些土地上的许多农民要么是穆斯林,要么更可能是采用了他们地主的宗教。 由于领土的原始性质,这些最初的定居者进入了人口大规模扩张的阶段,到现在孟加拉东部地区(孟加拉国)现在人口密度是西孟加拉邦(西孟加拉邦)的两倍。 这里的关键是,不需要大量的边缘泛灵论者,印度教徒或佛教徒农民大规模地conversion依。 相反,所有需要的只是适度的孟加拉国converted依农民,以成倍的人口增长,直到土地“被填满”。 (有趣的是,人们在美国和韩国人之间都发现了类似的模式。“核心”家园的宗教在宪法上与散居国外的人大不相同)
我认为这有说服力,主要有两个原因。 首先,彼得·贝尔伍德(Peter Bellwood) 第一农 记录尚未参与的人群的困难 集约 从农业转向这种方式。 至少可以追溯到莫卧儿时期,孟加拉是一个人口稠密的土地,仅仅因为总生产力,就可以从中获得大量租金。 如今,一个团结的孟加拉国将有240亿人口,使其成为世界第四人口大国,仅次于美国,仅次于印度尼西亚。 事后看来,我发现孟加拉东部的农民不太可能来自一直在练习的部落人民。 广泛 农业,但是被引进了新技术,比西方人早已习惯于集约化农业的残酷期望殖民化了“空”土地(事实上,孟加拉农民移居阿萨姆邦的部分原因是,尽管那里有土地过剩的感觉,尽管阿萨姆邦拥有30万居民)。 但是,殖民化的最初阶段将只需要相对较少的农民,并且可能表现出一些男性偏见。 因此,这可以解释孟加拉人中东方血统的很大一部分,因为在第一代中,孟加拉农民确实吸收了该地区的土著部落人民,无论该部落是 蒙达·桑塔尔斯(Munda Santhals) 或Tripura的藏缅亲戚。 与藏传缅甸人相比,孟加拉人数量众多,因此在任何模型中似乎都需要大量的基因流,因为这假定了这两组人在很长一段时间内的交流解释了人们发现非基因组的高比例的原因。 -南亚血统。 在 所有 印度只有10万讲藏缅语的人,而印度次大陆仅孟加拉语的240亿。
这在哪里离开我们? 根据我的收集,您可能不会进入第一轮结果 行动计划,但是如果您有23andMe个结果,但尚未将其发送到 扎克,并想进一步了解印度次大陆的历史遗传学, 您仍然可以参与其中! 有了我的父母,扎克现在拥有N = 2的孟加拉国。 希望能得到更多。 我们仍然需要来自印度中北部的样品。 旁遮普语的数量在5到10之间,泰米尔人的数量大约在5。 在不久的将来,我将从其他兄弟姐妹那里获得结果,并且我决定“升级”到V3芯片。 一旦出现,我将分阶段一些结果,并可能开始将自己与兄弟姐妹进行比较,“分阶段”结果,等等。
*美洲原住民是前哥伦布时期美国人的后裔,而南亚人(主要是亚洲人和欧洲少数人)的情况则相反。 这不仅仅是由于最近欧洲掺混物。 确切地说,尽管美洲印第安人与东亚有血缘关系,但这两个族群至少已有一万年的独特历史,而且可能更长。
**另外,有人说Noakhali的人狡猾狡猾,善于遵循 邮件 法律,而不是精神。 我只知道这一点,因为我小时候父亲的一位朋友,也来自孟加拉国,就抱怨说,诺卡里人彼此相识,而诺卡里人非常虔诚(他把妻子放在家里 面纱 当她从孟加拉国抵达时)要求其他人为他购买色情杂志。 他的理由是他不想被看到购买该杂志。 为好穆斯林购买这种物品是一种罪过。 后来,我的父亲和他的朋友(因身价从孟加拉国北部来)感到很同情,这是Noakhali人民的方式,在其中,您必须对自己有一些见识,以免他们为自己的利益谋取某种角度。 虔诚的非购房者以与其他孟加拉国人住在一起时不支付租金或迟交租金而臭名昭著,他始终强调他的宗教虔诚是最后偿还债务的保证。 他最终也发现了当时的移民法中的漏洞,相对容易地获得了绿卡,并且不需要赞助。 美国英语单词可能捕获了来自Noakhali的人如何居住的正确含义。 浮油.
 RSS
RSS


 就像“种群思维”一样,“树思维”对于理解进化生物学至关重要。 但这有问题。 首先,即使是在宏观进化尺度上,由于横向基因流动,无论是直接还是通过病毒介导,都严重违反了生命树各分支之间的分离。 当您细化到较细的系统发育粒时,就会遇到网状结构的问题,即将树转变为树状结构。 图形,就成为您必须集成到模型中的东西。 这是原因之一 混合树 这项技术被开发出来,使人们可以在微观进化规模上模拟跨树枝的基因流动。 但是还有另一个主要问题,那就是克莱因族。 TreeMix生成的“图形”仍然模拟脉冲混合事件。 但是,许多遗传变异是在以下情况下产生的: 距离隔离 动力学。 没有人口混合的奇异事件,而是每个实例都是连续混合过程的一部分。*这种情况的一个很好的例证是 环种.
就像“种群思维”一样,“树思维”对于理解进化生物学至关重要。 但这有问题。 首先,即使是在宏观进化尺度上,由于横向基因流动,无论是直接还是通过病毒介导,都严重违反了生命树各分支之间的分离。 当您细化到较细的系统发育粒时,就会遇到网状结构的问题,即将树转变为树状结构。 图形,就成为您必须集成到模型中的东西。 这是原因之一 混合树 这项技术被开发出来,使人们可以在微观进化规模上模拟跨树枝的基因流动。 但是还有另一个主要问题,那就是克莱因族。 TreeMix生成的“图形”仍然模拟脉冲混合事件。 但是,许多遗传变异是在以下情况下产生的: 距离隔离 动力学。 没有人口混合的奇异事件,而是每个实例都是连续混合过程的一部分。*这种情况的一个很好的例证是 环种.




















 在我
在我 



 到目前为止,我主要谈论基因。 但是就分类而言,基因没有什么神奇的。 生物人类学家使用
到目前为止,我主要谈论基因。 但是就分类而言,基因没有什么神奇的。 生物人类学家使用 
{kind=link}
{kind=link}